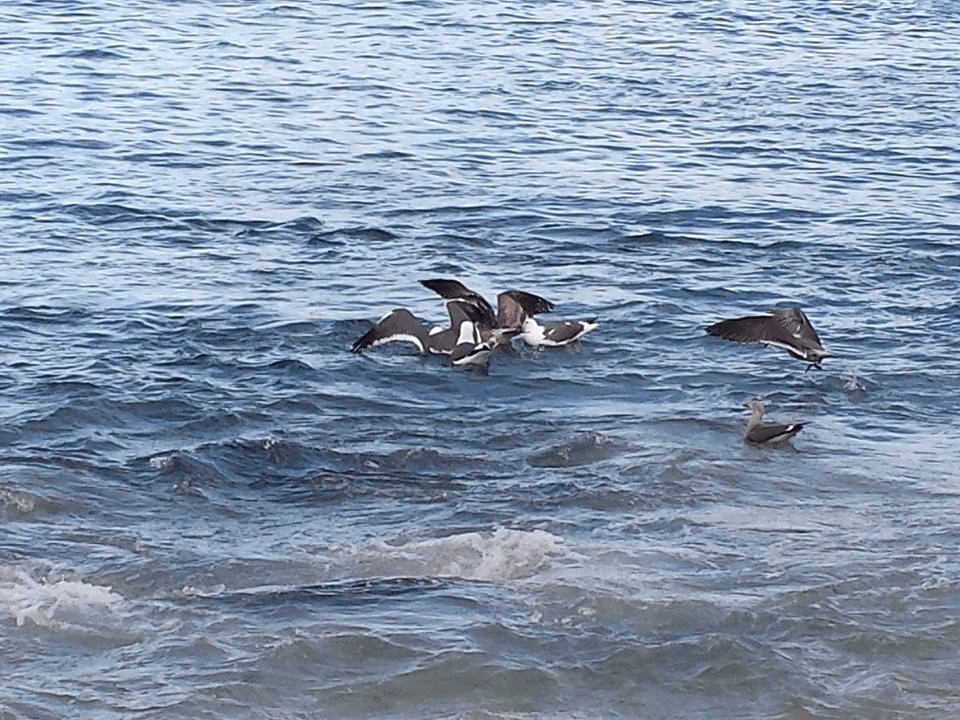On a late spring morning along Surf City, the first movement is often above the water, not within it. Brown pelicans travel low and steady just beyond the breakers, their wingtips nearly touching the surface as they follow a line that seems invisible from shore. Farther out, a group of terns holds in place against the wind, hovering, adjusting, then dropping sharply into the water before rising again. Closer to the sound side of Topsail Island, an osprey circles once, then folds into a dive toward a channel edge that looks, at first glance, no different than the water around it.
Nothing about these movements is random. They are responses to structure that exists beneath the surface—structure shaped by tide, wind, and the movement of other organisms. What appears as scattered bird activity is, in practice, a map of where the water is concentrating life.
For someone standing at the edge of it, that movement is one of the most accessible ways to read what cannot be seen directly.
What Birds Are Following Beneath the Surface
The birds that move along this stretch of coast are not searching broadly; they are tracking concentration. Along barrier island systems like those in Onslow County, physical processes—tidal exchange through inlets, wind-driven surface currents, and subtle differences in bottom shape—create zones where small fish, shrimp, and other prey accumulate (Peterson & Peterson, 1979; Piersma, 1997).
When the tide moves through places like New River Inlet, water does not flow evenly across the landscape. It accelerates through constrictions, slows along marsh edges, and bends around sandbars and channels. These shifts in speed and direction compress organisms into tighter spaces, particularly along boundaries where moving water meets something that resists it—an edge, a drop-off, or a change in depth (Wright et al., 1985).
Small schooling fish respond to that compression by tightening their formation. In doing so, they become more visible and more vulnerable. Larger fish—bluefish, Spanish mackerel, and juvenile coastal sharks—often move in from below, using that same concentration to feed. The pressure from below pushes prey upward, sometimes all the way to the surface.
Coastal birds feeding where prey has been concentrated near the surface along the breakers. | Image credit: A. Mitchell
What appears overhead depends on which part of that concentration each species is built to exploit.
Terns hovering and diving are often responding to prey that has been driven upward by predatory fish (Safina & Burger, 1985). Brown pelicans, which rely on plunge-diving, tend to follow more stable schools of fish that remain near the surface for longer periods (Shields, 2014). Ospreys, in contrast, depend on clear water and individual fish they can visually isolate, which is why their activity often aligns with calmer conditions and defined channel edges (Poole et al., 2002).
Each species is not simply feeding in the same place; each is reading a different layer of the same system.
When Surface Activity Signals Pressure Below
From the shoreline, bird activity can appear as isolated events—one dive, then another, then a sudden shift down the beach. Watched over time, a pattern emerges. A cluster of terns may concentrate in one location for several minutes, then disperse abruptly, reforming farther along the shoreline. Pelicans may align along a narrow band just beyond the breakers, following it as it drifts.
These shifts often reflect changes in how prey is being compressed and released beneath the surface. When predatory fish move through a bait school, the school tightens, rises, and becomes briefly accessible from above. When that pressure dissipates, the school spreads out again, and the birds move on.
This movement of energy—from smaller organisms to larger predators, and upward through the water column—is one visible expression of a trophic cascade. The term itself is often used to describe longer chains of ecological influence, but along the coast it can be observed in compressed moments, where the effects of predation become visible within seconds (Heithaus et al., 2008).
Birds do not initiate this process. They respond to it. Their presence marks where the system has already intensified.
Indicator Species at the Water’s Edge
From the beach, the difference is subtle. The water does not change color dramatically, and the waves continue to break as they did before. The level of activity shifts within that band—first visible in the air, then inferred below– marking places where the system has tightened, energy is moving through multiple layers at once, and the distance between surface and depth has, for a time, narrowed (Heithaus et al., 2008; Estes et al., 2011).
For someone entering the water, these differences in bird behavior can offer practical information, not in a predictive or absolute sense, but as indicators of what is happening just below the surface.
Brown pelicans traveling low in a consistent line often indicate schools of fish moving parallel to shore. Terns repeatedly diving in a tight area suggest smaller prey being pushed upward, frequently by larger fish feeding below. Ospreys focusing on a specific channel edge reflect clearer water and individual prey availability, rather than broad schooling events. Along the shoreline, shorebirds probing the sand at low tide are responding to invertebrates exposed by receding water, signaling a different layer of the system entirely—one tied to sediment and tidal timing rather than active predation (Colwell, 2010; Piersma, 1997).
None of these signals point directly to a specific species beneath the surface. What they indicate is concentration, and concentration is what draws larger predators closer to shore.
Along the coast of North Carolina, nearshore and juvenile shark presence is often associated with areas of high prey density, particularly where schooling fish aggregate (Heupel & Hueter, 2002). These conditions are not constant, and they shift with tide, temperature, and time of day. Birds make those shifts visible in real time.
At times, that activity stretches into lines that run the length of the breakers.
For someone stepping into the water, that narrowing matters. Not as a warning in the abstract, but as a recognition that the conditions supporting visible feeding above often extend below, linking organisms that are rarely seen together into the same moving structure.
Where the System Tightens
The patterns become easier to see near places where the water is forced to narrow, turn, or accelerate. The most consistent bird activity along this coast tends to occur where water movement is constrained and redirected. Inlets, marsh edges, sandbars, and the transitions between the Intracoastal Waterway and adjacent sounds create these zones (Wright et al., 1985).
At New River and its inlet, tidal flow compresses water into narrow channels before releasing it into broader areas, creating gradients in speed and depth. Along these gradients, prey accumulates, predators follow, and birds gather above.
These are not fixed points. As tide rises and falls, and as wind reshapes surface conditions, the locations of these compression zones shift. The birds move with them, tracing patterns that are constantly changing but not random.
For someone watching from shore, these movements can be read as lines, clusters, and absences—places where activity intensifies, and places where it suddenly drops away.
Standing Within It
Entering the water along this coast means stepping into a system already in motion. The surface may appear uniform, but the activity above it often reveals where that motion is focused.
Birds diving repeatedly in a confined area, or tracking a narrow band just beyond the breakers, indicate where prey is concentrated. Those same conditions are what draw larger predators into closer proximity to shore, not as an anomaly, but as part of the same process.
Watching the birds does not eliminate risk, and it does not provide certainty about what is beneath the surface. What it offers is context—a way to recognize when the water is more active, more compressed, and more connected across its layers.
What appears as feeding from above is part of a larger structure moving through the water. The birds do not create it, and they do not remain once it passes. They mark it, briefly, making visible what is otherwise difficult to see.
Bird movement along the shoreline often draws attention toward activity that remains unseen beneath the surface. | Image credit: A. Mitchell
References
Castro, J. I. (1993). The shark nursery of bulls Bay, South Carolina, with a review of the shark nurseries of the southeastern coast of the United States. Environmental Biology of Fishes, 38(1-3), 37-48. https://doi.org/10.1007/bf00842902
Colwell, M. A. (2010). Shorebird ecology, conservation, and management. University of California Press.
Estes, J. A., Terborgh, J., Brashares, J. S., Power, M. E., Berger, J., Bond, W. J., Carpenter, S. R., Essington, T. E., Holt, R. D., C. Jackson, J. B., Marquis, R. J., Oksanen, L., Oksanen, T., Paine, R. T., Pikitch, E. K., Ripple, W. J., Sandin, S. A., Scheffer, M., Schoener, T. W., & Wardle, D. A. (2011). Trophic downgrading of planet Earth. Science, 33(6040), 301-306. https://doi.org/10.1126/science.1205106
Heithaus, M. R., Frid, A., Wirsing, A. J., & Worm, B. (2008). Predicting ecological consequences of marine top predator declines. Trends in Ecology & Evolution, 23(4), 202-210. https://doi.org/10.1016/j.tree.2008.01.003
Heupel, M. R., & Hueter, R. E. (2002). Importance of prey density in relation to the movement patterns of juvenile blacktip sharks ( Carcharhinus limbatus ) within a coastal nursery area. Marine and Freshwater Research, 53(2), 543-550. https://doi.org/10.1071/mf01132
Piersma, T. (1997). Do global patterns of habitat use and migration strategies Co-evolve with relative investments in Immunocompetence due to spatial variation in parasite pressure? Oikos, 80(3), 623-631. https://doi.org/10.2307/3546640
Poole, A. F., Bierregaard, R. O., & Martell, M. S. (2002). Osprey (Pandion haliaetus). In The Birds of North America (1st ed.). Cornell Lab of Ornithology.
Safina, C., & Burger, J. (1985). Common tern foraging: Seasonal trends in prey fish densities and competition with bluefish. Ecology, 66(5), 1457-1463. https://doi.org/10.2307/1938008
Shields, M. (2014). Brown Pelican (Pelecanus occidentalis). In Birds of North America (1st ed.). Cornell Lab of Ornithology.
Wright, L., Short, A., & Green, M. (1985). Short-term changes in the morphodynamic states of beaches and surf zones: An empirical predictive model. Marine Geology, 62(3-4), 339-364. https://doi.org/10.1016/0025-3227(85)90123-9
There are mornings along the edges of the water in Onslow County when the surface looks still enough to trust.
The marsh grass has not yet reached its summer height. What stands there leaves more water exposed between the stems, and without sustained wind, the surface holds its shape. You can see farther into it now than you will in a few weeks, before suspended sediment and constant movement return it to opacity. The water carries less of the season, and because of that, more of what moves beneath it becomes visible—if you are willing to wait long enough to see the difference between movement and reflection.
This is when people begin to notice them again.
Not all at once. Not everywhere. Just a change that does not follow wind or tide. A line that holds where the rest of the surface releases. Something that holds its position in a system that is always adjusting.
An alligator does not arrive in that moment.
It becomes visible.
Alligator emerging from the mud. | Photo credit: Gilbert Grant, iNaturalist
Seasonal Absence Is Not Absence
Through winter, they remain within these same creeks, marsh edges, and quieter channels. What changes is not location, but how they occupy it. As temperatures fall, activity narrows. Movement slows, and the need for it slows with it. Energy is conserved, not spent. And the surface carries fewer signs of what lies beneath it. Individuals hold in deeper water or along softer margins where mud retains heat longer than the surrounding water column, remaining within conditions that allow them to persist without constant movement (Nifong et al., 2014; Rosenblatt & Heithaus, 2011).
The same stretch of water that in spring will hold a visible form can pass through winter without interruption, its stillness mistaken for absence.
But the system does not empty.
It compresses.
The System Wakes in Layers
By early spring, that compression begins to release—not all at once, but in layers that build on each other before they are recognized. Shallow water warms first, taking in solar heat more quickly than deeper channels. Along these edges, fish begin to hold longer. Movements that in winter passed through quickly begin to extend into areas that had remained quiet. Invertebrates return to the sediment surface, and the water column begins to carry more suspended life, even before it becomes visible as turbidity.
Birds respond to this before most other changes are noticed. Their movements tighten. Landings become more frequent, departures more abrupt. What they are tracking is not random. It is the redistribution of energy into places where it can be accessed.
The alligator moves within that shift.
Not as a trigger. Not as something layered on top. But as part of a system reorganizing itself across temperature, light, and movement at the same time.
Great blue heron and alligator are part of an interconnected system. | Photo credit: Audubon North Carolina
Reading What It Is Responding To
When one becomes visible along the edge of a creek or marsh, it is easy to reduce that moment to temperature alone. Warmer water allows for more activity.
But what draws it into that position is more specific than warmth.
It is the arrangement of prey.
Along the margins where water meets land, movement compresses. Fish traveling with the tide encounter shallow gradients that limit how long they can remain. Small mammals moving between marsh and upland must cross exposed edges. Birds landing to feed do so in places where depth and access align for only short intervals.
These are not isolated events. They are recurring patterns shaped by tidal cycles, substrate, and seasonal change.
The alligator positions itself within those patterns.
Its diet reflects that flexibility, spanning invertebrates, fish, birds, reptiles, and mammals depending on size and availability (Nifong, 2016). But the diet alone does not explain its placement. What matters is where energy becomes concentrated, even briefly.
That concentration is not constant. It forms and dissolves with tide, with light, with movement.
And the predator tracks that.
And what appears as a single movement—a fish turning, a bird lifting, something crossing the edge of the marsh—is part of a larger structure that holds only briefly before dissolving again.
The alligator does not respond to the individual movement.
It responds to the pattern that produces it.
Where Freshwater Meets Salt
These are not just places where water mixes.
They are places where movement is forced—and where that movement becomes available to something waiting at the top of it.
There are places along this coastline where those changes concentrate.
At the mouths of creeks, along the edges of the Intracoastal Waterway, and near the shifting bars of New River Inlet, the water does not settle into a single condition. Freshwater moves outward with tide and rainfall, meeting saltwater pressing back in with tidal exchange. The result is not a fixed boundary, but a gradient that shifts continuously—sometimes visible as a faint line, sometimes only detectable in how the surface moves differently from one side to the other.
This is where alligators are most often encountered—because this is where the system compresses into something they can use.
They are not marine animals. They do not possess the specialized salt glands that allow for extended life in high salinity environments. Over time, saltwater carries a physiological cost, requiring a return to freshwater to restore balance (Rosenblatt & Heithaus, 2011; Fujisaki et al., 2014).
But that limitation does not exclude them.
It defines how they move through them.
In these mixing zones, salinity is not constant. It rises and falls with tide, with rainfall, with wind direction. A location that carries higher salinity at one stage may shift toward fresher conditions hours later. What appears to be a boundary is, in practice, a moving field.
Within that field, movement compresses.
Fish traveling with the tide are funneled into narrower pathways. Shallow gradients limit how long they can remain in deeper water. Schools tighten. Individuals encounter edges that restrict escape. The system concentrates energy into space.
The predator does not need to range widely in these conditions.
It needs to hold where movement is forced.
And so it does.
An alligator near the tall grass near Marine Corps Air Station New River | Photo credit: Martin Egnash
At the Edge of the Open Water
There are moments when that pattern extends beyond the mixing zones, into places that appear, at first, outside of where an alligator belongs.
Along the shoreline, in the breaking waves where the ocean meets sand, one will sometimes appear—rising and falling with the swell, holding position just beyond where the water turns over onto the beach. It looks misplaced, as though it has moved beyond the system that defines it.
It has not.
The surf zone is one of the most compressed environments along the coast. Waves reduce depth, disrupt orientation, and concentrate movement into a narrow band where escape is limited. Fish pushed into breaking water lose some ability to maintain direction. Schools fragment. Individuals become briefly exposed in ways that do not occur in deeper, more stable water.
For a predator capable of stillness followed by short bursts of movement, that compression creates opportunity.
But the cost is higher.
Salinity is elevated. The water is in constant motion. There is no stable refuge within immediate reach. Time in this environment cannot be extended indefinitely.
And so it does not.
Movements into higher salinity water tend to be brief—extensions outward, followed by a return to freshwater or lower salinity conditions where balance can be restored (Nifong et al., 2014).
What appears as an anomaly is part of a larger pattern.
The predator crosses the boundary not to remain, but to use it, moving where the system briefly offers more than it costs.
The same forces that shape the marsh edge—compression, constraint, and brief exposure—are recreated here, just for a moment, in a different place.
An alligator rests at the ocean’s edge in North Topsail. | Photo credit: Fox8 Digital Desk
What Its Presence Changes
Most of what that presence changes cannot be seen when it is observed.
Long before any direct interaction occurs, it is already altering how other organisms use space.
Fish moving along the edge do not simply pass through. They adjust their depth, their speed, the amount of time they remain exposed. Birds land with shorter intervals between contact and departure. Mammals approaching the water shift their paths or their timing. These changes are not dramatic in isolation. But they are continuous.
Over time, they accumulate into structure—the kind that determines who feeds, where they feed, and how long they remain.
The influence of a predator at this level extends beyond what it consumes. It shapes behavior across multiple species, redistributing where and how energy moves through the system. The possibility of predation—present even when not observed—alters interactions in ways that regulate access to habitat and resources (Heithaus et al., 2008; Ripple et al., 2014; Estes et al., 2011).
What holds the system in place is not removal alone.
It is pressure.
What is being shaped is not just movement, but access—and access is what determines how energy moves through the system.
More Than Predation
The influence of the alligator does not end with what it hunts, but extends beyond those interactions.
As it moves through shallow systems, it disturbs sediment, creating depressions and pathways that alter how water is retained and how nutrients are redistributed. These small changes in physical structure create conditions that other species use—temporary refuges, feeding areas, and zones where organic material accumulates (Eversole et al., 2018; Subalusky et al., 2009).
In wetland systems, these disturbances have been linked to broader effects, including nutrient cycling and carbon storage, where the presence of large predators contributes to the retention of organic material within the system rather than its export (Murray et al., 2025; Atwood et al., 2015).
These processes do not occur in isolation.
They intersect with the same patterns of movement, feeding, and behavior that define the system at larger scales.
Seeing the Surface, Reading the System
When one becomes visible along the surface, it is easy to treat the moment as singular.
A sighting. An encounter. Something separate from everything around it.
But that form at the surface is supported by layers extending beyond what can be seen.
It reflects water temperatures crossing into ranges that support sustained activity. It reflects prey moving into positions where access becomes possible. It reflects a system where behavior is still shaped by the presence of something at the top.
The alligator is not an interruption to that system.
It is an expression of it.
What Becomes Visible
Seeing one does not indicate that something has entered the water.
It indicates that enough beneath the surface is functioning to hold it.
Not in a static sense. Not as balance in the way it is often described. But as a set of interactions that remain connected—movement, response, pressure—each shaping the others even when they are not directly observed.
What becomes visible at the surface is only a fraction of that structure.
But it is enough to know that the rest is still in place.
An alligator in Onslow County sits at the edge of the saltmarsh. |Photo credit: Gilbert Grant, iNaturalist
When That Pressure Is Reduced
If that pressure is reduced, the system does not leave an obvious gap.
It shifts.
Movements that were once constrained begin to extend. Species that passed quickly through exposed areas begin to remain longer. Edges that functioned as transition zones become used differently—not because the physical environment has changed, but because the conditions that shaped behavior within it have relaxed.
Mid-level predators expand their activity under these conditions, increasing their access to prey and space when not constrained from above (Nifong et al., 2013).
The change is subtle.
It appears in how long something stays. In how often it returns. In where it lingers. In how quietly the structure of behavior begins to loosen.
The food web and trophic cascade of the American alligator in the Florida Everglades.
A System Written Into Temperature
There is another layer to this that does not show itself at the surface.
The structure of that presence is set years earlier, in a place that can be overlooked when standing at the water’s edge. Along the margins of marsh and wetland, slightly above the reach of regular water movement, nests are built from vegetation and sediment, forming mounds that hold heat as they decompose.
Within those mounds, temperature determines something that will not be visible for much later.
Sex is not fixed at fertilization. It emerges during incubation, shaped by the thermal conditions held within the nest. A difference of only a few degrees is enough to shift the outcome, producing more males or more females depending on where within that range the nest remains (Lang & Andrews, 1994; Janzen, 1994).
Under variable conditions—differences in shading, rainfall, timing, and placement—those outcomes are distributed across the landscape. Some nests produce more females, others more males. That variability holds the population in a form that can sustain itself over time.
When conditions become more consistent, that variation narrows.
Warmer nights hold heat longer within the nest. Seasonal transitions extend. The range of outcomes compresses. What was once distributed begins to align.
And that alignment carries forward into the structure of the population—into how individuals occupy space, into how pressure is applied across the system, into what will eventually be visible at the surface.
Alligator eggs hatch after 65 days of incubation in the fall. The babies will chirp to alert their mom, who then digs out the nest while the babies use their egg tooth to hatch from their eggs. Their mom will then safely carry them to the water.
Where the Next Generation Is Set
The placement of those nests depends on something even more constrained.
A narrow band of land that remains above water just long enough to hold them.
That band is not fixed.
It shifts with tide, with rainfall, with the gradual reworking of shoreline that occurs across seasons and years. With rising sea levels, water reaches farther into areas that once remained above it. Flooding becomes more frequent, not always through singular events, but through repeated intrusions that saturate and destabilize what had previously held (Joanen & McNease, 1989; Sweet et al., 2022).
Human alteration compresses this space further.
Hardened shorelines, dredging, and development reduce the gradual transition between land and water. Where there was once a slope capable of holding multiple elevations, there becomes a defined edge. That edge does not provide the same range of conditions required for successful nesting.
The number of suitable sites decreases.
More importantly, the variability between them narrows.
And with that, the system loses one of the mechanisms that allowed it to absorb change.
Alligator on her nest that can hold up to 60 eggs. | Photo credit: National Park Service (NPS)
What Its Presence Means
When an alligator becomes visible along the surface, it reflects conditions that have aligned across multiple layers.
Temperature has reached a range that supports activity. Prey has moved into positions where access becomes possible. Behavioral pressure remains in place across the system. Reproduction has held across enough years, in enough suitable places, to sustain what is now present.
What is seen at the surface is not separate from them.
It is supported by them.
Seeing one does not signal that something has entered the water.
It signals that enough of what lies beneath it—movement, pressure, response, and continuity—remains intact.
And that—even when most of it is not visible—the system is still holding together.
And that is what becomes visible—just long enough to be seen, before the system closes back over it again.
The system does not end at the water’s edge.
Epilogue: Chicken Nugget
We came across him along the New River, near the courthouse in Jacksonville.
We were there to clear what had been left behind—fishing line caught along the walkways, hooks, and the overflow from a trash can that had spilled out onto the edge. Fast food containers, grocery store chicken trays, scattered along the bank. The signs were clear enough. People had been there for a while—crabbing, fishing, eating, leaving what remained.
He was directly below us.
Small enough to miss at first. Still enough to blend into the water until you stopped looking for movement and started noticing what held its position.
A juvenile alligator, watching.
He stayed there while we worked, then slipped beneath the surface and crossed the small bay. On the opposite side, someone tossed a piece of food into the water. He surfaced almost immediately, took it, and remained.
Waiting.
I came back later and stayed longer.
The pattern repeated. He would disappear until footsteps approached, then return to the same place along the edge. Holding position. Watching. Waiting for something to fall.
No fishermen or crabbers passed through while I was there, but the behavior was consistent with what happens when food becomes predictable. Bait, catch, scraps—anything that can be taken without the cost of searching or pursuing.
Energy, without effort.
It is easy to see something like that and respond to what it looks like in that moment. A small animal. Still. Attentive. Something that feels close enough to interact with.
But what is being shaped there is not just a single interaction.
It is behavior.
A shift away from the conditions that formed it—toward something more efficient, more immediate, and less stable over time. The system that once required movement, patience, and response begins to narrow into expectation.
And expectation changes how an animal uses space.
What happens when that animal is no longer small is not a separate question.
It is the continuation of the same pattern.
Alligators do not forget where food has been easy to obtain. They return to it. They hold in those places. They begin to associate presence—human presence—with opportunity.
What begins as something that feels harmless becomes something that alters how the system functions around it.
Not just for the animal, but for everything that responds to it.
There are instincts at work here that were shaped long before any walkway, any dock, any place where food might be dropped from above. Those instincts are not just about survival in isolation. They are part of how pressure is applied, how movement is shaped, how the system holds.
When those instincts are replaced with something easier, the effect does not remain contained.
It carries outward.
He stayed there while I watched. Returning to the same place. Holding the same position. Waiting for something to fall.
There is a kind of kindness in wanting to give something to an animal like that.
But there is another kind in leaving it as it is.
Not interrupting the conditions that shape it. Not narrowing what it has learned to expect. Not replacing a system built on movement and response with one built on waiting.
Let it remember the water as it is.
And you, only as something that passed through it.
We affectionately named this juvenile alligator in the New River in Jacksonville, NC “Chicken Nugget” for all of the chicken nugget boxes left behind on the walkway from an overflowing trash can. | Photo credit: A. Mitchell
References
Atwood, T. B., Connolly, R. M., Ritchie, E. G., Lovelock, C. E., Heithaus, M. R., Hays, G. C., Fourqurean, J. W., & Macreadie, P. I. (2015). Predators help protect carbon stocks in blue carbon ecosystems. Nature Climate Change, 5(12), 1038-1045. https://doi.org/10.1038/nclimate2763
Estes, J. A., Terbough, J., Brashares, J. S., Power, M. E., Berger, J., Bond, W. J., Carpenter, S. R., Essington, T. E., Holt, R. D., & Wardle, D. A. (2011). Trophic Downgrading of Planet Earth. Science, 333(604), 301-306. https://www.science.org/doi/abs/10.1126/science.1205106
Fujisaki, I., Hart, K. M., Mazzotti, F. J., Cherkiss, M. S., Sartain, A. R., Jeffery, B. M., Beauchamp, J. S., & Denton, M. (2014). Home range and movements of American alligators (Alligator mississippiensis) in an Estuary habitat. Animal Biotelemetry, 2(1), 8. https://doi.org/10.1186/2050-3385-2-8
Heithaus, M. R., Frid, A., Wirsing, A. J., & Worm, B. (2008). Predicting ecological consequences of marine top predator declines. Trends in Ecology & Evolution, 23(4), 202-210. https://doi.org/10.1016/j.tree.2008.01.003
Janzen, F. J. (1994). Climate change and temperature-dependent sex determination in reptiles. PNAS, 91(16), 7487-7490. https://doi.org/10.1073/pnas.91.16.7487
Joanen, T., & McNease, L. L. (1989). Ecology and physiology of nesting and early development of the American alligator. American Zoologist, 29(3), 987-998. https://doi.org/10.1093/icb/29.3.987
Lang, J. W., & Andrews, H. V. (1994). Temperature‐dependent sex determination in crocodilians. Journal of Experimental Zoology, 270(1), 28-44. https://doi.org/10.1002/jez.1402700105
Nifong, J. C. (2016). Living on the edge: Trophic ecology of alligator mississippiensis (American alligator) with access to a shallow estuarine impoundment. Bulletin of the Florida Museum of Natural History, 54(2), 13-49. https://doi.org/10.58782/flmnh.xkdw7119
Nifong, J. C., Nifong, R. L., Silliman, B. R., Lowers, R. H., Guillette, L. J., Ferguson, J. M., Welsh, M., Abernathy, K., & Marshall, G. (2014). Animal-borne imaging reveals novel insights into the foraging behaviors and Diel activity of a large-bodied APEX predator, the American alligator (Alligator mississippiensis). PLoS ONE, 9(1), e83953. https://doi.org/10.1371/journal.pone.0083953
Nifong, J. C., & Silliman, B. R. (2013). Impacts of a large-bodied, APEX predator (Alligator mississippiensis Daudin 1801) on salt marsh food webs. Journal of Experimental Marine Biology and Ecology, 440, 185-191. https://doi.org/10.1016/j.jembe.2013.01.002
Ripple, W. J., Estes, J. A., Beschta, R. L., Wilmers, C. C., Ritchie, E. G., Hebblewhite, M., Berger, J., Elmhagen, B., Letnic, M., Nelson, M. P., Schmitz, O. J., Smith, D. W., Wallach, A. D., & Wirsing, A. J. (2014). Status and ecological effects of the world’s largest carnivores. Science, 343(6167). https://doi.org/10.1126/science.1241484
Rosenblatt, A. E., & Heithaus, M. R. (2011). Does variation in movement tactics and trophic interactions among American alligators create habitat linkages? Journal of Animal Ecology, 80(4), 786-798. https://doi.org/10.1111/j.1365-2656.2011.01830.x
Sweet, W. V., Hamlington, B. D., Kopp, R. E., Weaver, C. P., Barnard, P. L., Bekaert, D., Brooks, W., Craghan, M., Dusek, G., Frederickse, T., Garner, G., Genz, A. S., Krasting, J. P., Larour, E., Marcy, D., Marra, J. J., Obeysekera, J., Osler, M., Pendleton, M., … Zuzak, C. (2022). Global and regional sea level rise scenarios for the United States: Updated mean projections and extreme water level probabilities along U.S. coastlines (Technical Report NOS 01). National Oceanic and Atmospheric Administration, National Ocean Service. https://earth.gov/sealevel/us/internal_resources/756/noaa-nos-techrpt01-global-regional-SLR-scenarios-US.pdf
A breath breaks the air first — a soft exhale that sounds almost human — and then a dorsal fin lifts from the channel like a line drawn through moving water. The tide is falling. Gulls hover over the seam where current tightens. Fishermen pause mid-cast because everyone knows the rhythm: if the dolphins are working the edge, the fish are already gathering.
These encounters feel spontaneous, but they are not accidents. The dolphins that surface beside our piers, marsh creeks, and inlets are not anonymous travelers passing through. Many bottlenose dolphins show long-term site fidelity and structured community patterns in estuarine systems, returning to the same places across years (Urian et al., 2009; Wells, 2014). To live on this shoreline is to share space with minds moving just below the surface — residents of the tidal edge.
Who they are: a coastal population
The dolphins most frequently seen along Onslow County’s waters are common bottlenose dolphins (Tursiops truncatus), a species whose “coastal” lives can look very different from “offshore” lives. Across the western North Atlantic, genetic studies show fine-scale population structure that can separate dolphins using nearshore coastal waters from dolphins using inshore estuarine waters (Rosel et al., 2009). More broadly, integrative work continues to support meaningful coastal vs offshore divergence in the region (Costa et al., 2022).
In estuaries, photo-identification research (matching dorsal-fin markings) repeatedly shows that bottlenose dolphins can form discrete social communities with limited spatial overlap — a pattern consistent with long-term residency and local familiarity (Urian et al., 2009). In practical terms, the dolphin a child watches from a dock in spring may be seen again the following winter, and again the next year: not a rumor, but a biological possibility supported by long-term studies of resident dolphins elsewhere on the coast (Wells, 2014).
Photo-identification doesn’t always rely solely on human matching of fin shapes; new tools such as machine learning are being developed to improve accuracy in identifying individual dolphins and whales in the wild. For example, researchers in Hawaii are using advanced algorithms to distinguish individuals from large photo libraries of dorsal fins. As technology improves, methods like photo-ID only get more reliable — which means studies of habitat overlap and seasonal return become more precise over time.
An inside look at how scientists “read” dorsal fin shapes and markings to track the same dolphins over time.
Reading the geometry of the estuary
Dolphins do not simply occupy estuaries; they interpret them.
Tidal channels function as moving architecture. Falling tides compress fish schools toward narrowing exits. Sandbars redirect flow into faster seams. Marsh edges trap prey against shallow gradients. Dolphins exploit these features with precision, repeatedly targeting conditions that make prey capture more efficient (Barros & Wells, 1998; Torres & Read, 2009).
This is one reason dolphins so often appear where the water “looks alive” — at convergence lines, inlet throats, and channel bends. In Florida Bay, for example, foraging tactics are mapped onto habitat features that define where dolphins have spent their time, thus turning behavior into geography (Torres & Read, 2009). What seems like play from shore can be highly strategic predation.
Bottlenose dolphins breaching off Seaview Pier, N. Topsail Beach, North Carolina. The arc of the body and column spray reflect the mechanics of propulsion – force directed through the tail, momentum carried into the air. | Photo credit: Howard Crumpler Photography, 2026
Reader Question:
Why do dolphins seem more active on rainy or overcast days?
Weather, light, and the illusion of play
You may notice that dolphins seem especially active on overcast or rainy days — surfacing more frequently, breaching, or moving in tight arcs through wind-rippled water. It can look like preference, even mood. But dolphins are responding less to cloud cover than to what cloud cover does to the water.
When the sky darkens, baitfish don’t stay arranged the same way. They may bunch together or rise toward the surface. For a predator already working those upper layers, that shift can make hunting more efficient (Benoit-Bird & Au, 2003). Wind and rain can also stir the surface and cloud the water, changing who sees whom first (De Robertis et al., 2003).
There is also a perceptual component. Overcast skies reduce glare, making dorsal fins and splashes easier for human observers to detect. Wind-textured water highlights movement. What appears to be “more play” may sometimes be improved visibility — a reminder that observer experience and animal behavior are not always the same phenomenon.
In short, dolphins are responding to ecological conditions. The weather alters the water; the water alters the fish.
Two bottlenose dolphins break the surface beneath the gray horizon off Surf City, North Carolina. Overcast light and wind-roughened water can change how fish move – and how easily we notice the dolphins following them. | Photo credit: Johnny Provost, Jr., 2025
Communication and social intelligence
Bottlenose dolphins have been studied for decades not just because they are charismatic, but because their social lives depend on constant communication in a shifting, three-dimensional world. One of the strongest findings to emerge from that research is the existence of signature whistles — individually distinctive call types that function as learned identity signals, something very much like the individual name a dolphin goes by within its community (Janik & Sayigh, 2013).
Social learning runs just as deep. Some dolphin foraging habits spread from one animal to another rather than through genetics — passed along socially, a rare pattern among nonhuman species (Krützen et al., 2005). Mothers and calves stay together for years, giving calves time to learn not just how to hunt, but where — which channels to follow, which bends of water hold fish (Wells, 2014).
In some populations elsewhere in the world, dolphins even use tools — carrying marine sponges on their rostrums while foraging or trapping fish inside empty shells — behaviors that are socially learned and culturally transmitted (Krützen et al., 2005).
That learning shapes how dolphins fit into the estuary. In many tidal systems they sit near the top of the local food web, influencing the fish communities beneath them. Yet beyond those protected waters, they are not beyond risk. Large sharks prey on dolphins, placing them within a broader coastal hierarchy where even predators can become prey (Heithaus, 2001). The role shifts with scale. The ecology remains layered.
Two bottlenose dolphins surfacing together off Seaview Pier, N. Topsail Beach, North Carolina. Close positioning and timing are hallmarks of the complex social bonds that define dolphin societies. | Photo credit: Howard Crumpler Photography, 2026
Dolphins are not guardians
Popular culture has assigned dolphins a role they never chose: protector. People repeat a comforting shoreline myth — “If you’re scared of sharks, find the dolphins; they’ll protect you.” But that story is not grounded in how dolphins behave in the wild.
Bottlenose dolphins are powerful predators. They compete, establish dominance hierarchies, and can deliver forceful blows when defending calves or asserting space. Dolphin–shark interactions occur, but they are not “rescue missions” staged for humans; they are ecological encounters shaped by risk, competition, and opportunity (Heithaus, 2001).
Wild dolphins are also capable of injuring people. Research examining human–dolphin interactions show that close approaches — and especially feeding wild dolphins — increase the likelihood of risky contact and harmful outcomes for both dolphins and people (Cunningham-Smith et al., 2006; Vail, 2016). Over time, those interactions leave visible consequences. Long-term data from Sarasota Bay show that dolphins who have learned to associate people with food are more likely to carry injuries linked to boats and fishing gear (Christiansen et al., 2016).
The danger is not that dolphins are “evil.” The danger is assuming they share human intentions.
Swimming near a pod does not create a protective shield. Dolphins are not lifeguards. They are wild animals navigating their own priorities in a shared environment. Respecting that boundary is what allows coexistence.
A bottlenose dolphin pursuing prey near a recreational vessel in a waterway in Surf City, North Carolina. Foraging behavior can bring dolphins into close proximity with boats – not as companions, but as active predators focused on fish. | Video credit: Cynthia Dirosse, 2024
Winter dolphins
A persistent assumption is that dolphins vanish when the water cools. In reality, seasonal distribution can be more nuanced — changing with prey, temperature, and coastal movement patterns rather than following a simple on/off presence.
Along the mid-Atlantic coast, research shows that bottlenose dolphins shift their movements with the seasons, appearing in different areas at different times of year (Torres et al., 2005). Studies focused on estuarine dolphins in southern North Carolina document similar seasonal patterns closer to home (Silva et al., 2020). From shore, those changes can look like disappearance. But winter quiet does not always mean absence. It may simply mean dolphins are working deeper channels or less visible pathways beyond the easy reach of our eyes.
The estuary in winter is quieter, but not empty.
Dorsal fins in winter light off Surf City, North Carolina. Dolphins may appear less active this time of year, but changes in light, water depth, and travel corridors often influence what we notice from shore. | Photo credit: Surf City Parks, Recreation, and Tourism, 2017
Living beside them
Living near dolphins is a privilege — and it places us within the same waters they navigate. Vessel traffic, fishing gear, and repeated close approaches can shape the lives of animals that live for decades and raise calves slowly (Wells, 2014). Studies of dolphins that have been fed or closely approached by people show that these interactions can shift behavior, making dolphins more likely to approach boats and increasing the risk of injury and conflict (Vail, 2016). Distance, in that sense, preserves the patterns people come to watch.
The presence of dolphins is not guaranteed. It is a sign that the system still functions — prey, water quality, shoreline structure, and the complex social knowledge dolphins carry from year to year. As long-lived predators near the top of the food web, they are indicator species, reflecting the condition of the waters they inhabit — estuary, inlet, and nearshore coast alike.
And so when a dorsal fin rises beyond the channel markers, it means more than a moment of spectacle. It means the currents are still working, the fish are still moving, and the layered relationships that shape this shoreline are still holding.
There is always more to learn about dolphins than fits in a single post. For those who’d like to go further, this episode of the All Creatures Podcast offers a thoughtful exploration of their biology and behavior.
References
Barros, N. B., Wells, R. S., & Barros, N. B. (1998). Prey and feeding patterns of resident bottlenose dolphins (Tursiops truncatus) in Sarasota Bay, Florida. Journal of Mammalogy, 79(3), 1045. https://doi.org/10.2307/1383114
Benoit-Bird, K. J., & Au, W. W. (2003). Prey dynamics affect foraging by a pelagic predator (Stenella longirostris) over a range of spatial and temporal scales. Behavioral Ecology and Sociobiology, 53(6), 364-373. https://doi.org/10.1007/s00265-003-0585-4
Christiansen, F., McHugh, K. A., Bejder, L., Siegal, E. M., Lusseau, D., McCabe, E. B., Lovewell, G., & Wells, R. S. (2016). Food provisioning increases the risk of injury in a long-lived marine top predator. Royal Society Open Science, 3(12), 160560. https://doi.org/10.1098/rsos.160560
Costa, A. P., Mcfee, W., Wilcox, L. A., Archer, F. I., & Rosel, P. E. (2022). The common bottlenose dolphin (Tursiops truncatus) ecotypes of the western North Atlantic revisited: An integrative taxonomic investigation supports the presence of distinct species. Zoological Journal of the Linnean Society, 196(4), 1608-1636. https://doi.org/10.1093/zoolinnean/zlac025
Cunningham-Smith, P., Colbert, D. E., Wells, R. S., & Speakman, T. (2006). Evaluation of human interactions with a provisioned wild bottlenose dolphin (<I>Tursiops truncatus</I>) near Sarasota Bay, Florida, and efforts to curtail the interactions. Aquatic Mammals, 32(3), 346-356. https://doi.org/10.1578/am.32.3.2006.346
De Robertis, A., Ryer, C. H., Veloza, A., & Brodeur, R. D. (2003). Differential effects of turbidity on prey consumption of piscivorous and planktivorous fish. Canadian Journal of Fisheries and Aquatic Sciences, 60(12), 1517-1526. https://doi.org/10.1139/f03-123
Heithaus, M. R. (2001). Shark attacks on bottlenose dolphins (TURSIOPS ADUNCUS) in Shark Bay, Western Australia: Attack rate, bite scar frequencies, and attack seasonality. Marine Mammal Science, 17(3), 526-539. https://doi.org/10.1111/j.1748-7692.2001.tb01002.x
Janik, V. M., & Sayigh, L. S. (2013). Communication in bottlenose dolphins: 50 years of signature whistle research. Journal of Comparative Physiology A, 199(6), 479-489. https://doi.org/10.1007/s00359-013-0817-7
Krützen, M., Mann, J., Heithaus, M. R., Connor, R. C., Bejder, L., & Sherwin, W. B. (2005). Cultural transmission of tool use in bottlenose dolphins. Proceedings of the National Academy of Sciences, 102(25), 8939-8943. https://doi.org/10.1073/pnas.0500232102
Rosel, P. E., Hansen, L., & Hohn, A. A. (2009). Restricted dispersal in a continuously distributed marine species: Common bottlenose dolphinsTursiops truncatusin coastal waters of the western North Atlantic. Molecular Ecology, 18(24), 5030-5045. https://doi.org/10.1111/j.1365-294x.2009.04413.x
Silva, D. (2020). Abundance and seasonal distribution of the southern North Carolina estuarine system stock (USA) of common bottlenose dolphins (Tursiops truncatus). IWC Journal of Cetacean Research and Management, 21(1), 33-43. https://doi.org/10.47536/jcrm.v21i1.175
Torres, L. G., McLellan, W. A., Meagher, E., & Pabst, D. A. (2023). Seasonal distribution and relative abundance of bottlenose dolphins, Tursiops truncatus, along the US Mid-Atlantic coast. J. Cetacean Res. Manage, 7(2), 153-161. https://doi.org/10.47536/jcrm.v7i2.748
Torres, L. G., & Read, A. J. (2009). Where to catch a fish? The influence of foraging tactics on the ecology of bottlenose dolphins (Tursiops truncatus) in Florida Bay, Florida. Marine Mammal Science, 25(4), 797-815. https://doi.org/10.1111/j.1748-7692.2009.00297.x
Urian, K. W., Hofmann, S., Wells, R. S., & Read, A. J. (2009). Fine‐scale population structure of bottlenose dolphins (Tursiops truncatus) in Tampa Bay, Florida. Marine Mammal Science, 25(3), 619-638. https://doi.org/10.1111/j.1748-7692.2009.00284.x
Vail, C. S. (2016). An overview of increasing incidents of bottlenose dolphin harassment in the Gulf of Mexico and possible solutions. Frontiers in Marine Science, 3. https://doi.org/10.3389/fmars.2016.00110
Wells, R. S. (2013). Social structure and life history of bottlenose dolphins near Sarasota Bay, Florida: Insights from four decades and five generations. Primatology Monographs, 149-172.