In the last few weeks, the water along the edges of Onslow County has felt different.
Not because the water itself has changed—but because something beneath it has become harder to ignore.
Schools of cownose ray (Rhinoptera bonasus) move just below the surface nearshore, their wingbeats lifting faint clouds from the bottom as they pass. In the soundside shallows, where the water thins over sand and mud, Atlantic stingray (Hypanus sabinus) settle into the substrate, half-buried and nearly invisible until a step comes too close and the outline breaks.
People are seeing them more often now—but they’re also reacting to them.
A pause mid-step in shallow water. A quick shift backward when something moves. Fishermen lifting a line and stopping for a second longer than usual—not what they expected to find.
There is awe in it.
And sometimes hesitation.
Because the same thing that makes them easy to notice now also makes them easy to miss.
The question follows quickly:
Are there more of them this year?
Maybe.
But that question lingers longer than the answer.
Cownose rays migrating in Swansboro, NC. | Image credit: Pogie’s Academy
What Brings Them Here
As spring settles in along the North Carolina coast, the system begins to reorganize.
Water temperatures rise, and with that rise comes a shift in metabolism. Rays—like many coastal species—become more active as conditions move into a narrower range that supports feeding and movement (Smith & Merriner, 1987; Schwartz & Dahlberg, 1978).
For cownose rays, this seasonal transition includes a northward migration along the Atlantic coast, bringing large groups into nearshore and estuarine waters (Smith & Merriner, 1987).
Large groups of cownose rays like these move north along our coast each season, arriving together in shallow water. | Image credit: Vidyacharan A. Alchi
But movement alone does not explain what people are seeing.
What matters is where that movement meets the structure of the environment.
The water does not always look the same—some days it is flat and clear enough to see straight to the bottom, and other days the slightest movement turns it cloudy, changing what can be seen and what remains hidden (Peterson et al., 2001).
And beneath all of it is food.
Cownose rays move through the shallows, sweeping across the bottom and disrupting what lies beneath them, crushing clams, oysters, and other shelled invertebrates with broad, flattened tooth plates (Collins et al., 2007; Fisher, 2010).
Atlantic stingrays hold low against the bottom, burying into the sand as they feed and working within the sediment itself—not moving across it—uncovering and drawing in small invertebrates hidden below (Snelson et al., 1988; Schwartz & Dahlberg, 1978).
Atlantic stingrays hold close to the bottom, often blending in until something shifts and gives them away. | Image credit: Andy Murch
Where prey is accessible, rays follow.
Where prey is concentrated in shallow, warming water, rays do not just pass through—they stay, turn, feed, and linger.
And in doing so, they cross into the same narrow band of space where people enter the water (Bangley et al., 2018).
They are not simply “here more.”
They are here in ways—and in places—that make them visible.
What Happens When They Feed
When a ray feeds, the bottom does not remain the same.
A cownose ray moving across a flat is not just searching—it is actively restructuring the surface beneath it. As it passes, the bottom is turned over behind it, patches of sand and mud disturbed where clams and other buried life have just been uncovered and crushed (Peterson et al., 2001; Smith & Merriner, 1985).
Feeding pits left behind by rays. Easy to mistake for crab holes at first—until you start to recognize the pattern and what’s actually shaping the bottom. | Image credit: Giaroli et al., 2024
Atlantic stingrays leave a different kind of trace. Where they settle, the surface shifts more subtly—small depressions, softened patches, places where the sediment has been worked rather than overturned, as buried invertebrates are uncovered and drawn in (Snelson et al., 1988; Schwartz & Dahlberg, 1978).
This is bioturbation—the bottom being reworked by the animals moving through it and within it (Thrush & Dayton, 2002).
As they feed, the bottom lifts into the water—fine particles rising and hanging there, turning clear water slightly cloudy (Thrush & Dayton, 2002).
Oysters and clams quietly filter the water as they feed, and when their numbers shift—even in small areas—the water and everything moving through it begins to change with them (Newell, 2004; zu Ermgassen et al., 2013).
In places where rays have been feeding, those filtering communities can be reduced or redistributed (Peterson et al., 2001).
Not removed entirely—but changed.
And that change does not stay in one place.
It moves outward, carried in the way the water looks, the way it settles, and what it can hold.
Layers of the Food Web
Rays do not sit at the top of the system, and they are not at the bottom of it.
As mesopredators, they feed on what is buried in the sediment, but they are also available to what moves through the water above. That position—between—links parts of the system that do not often meet directly (Myers et al., 2007; Heithaus et al., 2008).
What they do in that space matters.
As cownose rays move through andAtlantic stingrays work within the bottom, they are not just feeding—they are shaping what persists there. Clams, oysters, and other invertebrates do not simply accumulate unchecked. Their numbers are reduced, redistributed, and in some places kept from becoming dominant (Peterson et al., 2001).
Movement like this doesn’t stay in one place for long.
That pressure shapes the bottom itself.
Bivalves filter the water. Invertebrates stabilize sediment. When their abundance shifts, the system responds—sometimes toward clearer water, sometimes toward more suspended material, depending on what remains and where (Newell, 2004; zu Ermgassen et al., 2013).
Rays do not create those conditions alone—but they influence which direction the system moves.
At the same time, they carry that energy upward.
Juvenile sharks moving through these shallow waters encounter not just prey, but a system already in motion—areas where the bottom has been disturbed, where feeding has recently occurred, where something has been uncovered or displaced (Bangley et al., 2018).
And in some cases, the rays themselves become part of that exchange.
This is what it means to sit in the middle.
Not just connecting layers—but regulating how energy and movement pass between them.
If that middle shifts, the balance does not disappear.
It changes direction.
Why It Feels Sudden
There is a moment, standing in shallow water, when the bottom stops feeling like something you can trust.
What looked like sand shifts. What felt still is no longer still.
Sometimes you notice it in time—a shape lifting away, a shadow moving just beneath the surface. A plume of fine sediment rising to the surface under a paddleboard with a trail following it.
The moment when the bottom stops looking empty. | Image credit: iStock
Sometimes you don’t.
A step comes down where something is already settled. Hidden in the sand. Working within it.
The reaction is immediate. Surprise first. Then pain. Then the realization of what was there all along.
But what you are stepping into is not a single event.
It is a convergence.
Water temperatures have risen, bringing rays into the shallows as they feed and move through these systems (Smith & Merriner, 1987; Schwartz & Dahlberg, 1978).
Tides narrow the space, concentrating movement into a thinner band of water.
The bottom has already been worked—turned by cownose rays moving through, disturbed by Atlantic stingrays holding within it.
And at the same time, people have returned to the water.
For a brief window, all of it overlaps.
Not more. But more visible.
It feels sudden because you are standing at the point where all of these things meet.
And for a moment, the system lets you see it.
References
Bangley, C. W., Paramore, L., Dedman, S., & Rulifson, R. A. (2018). Delineation and mapping of coastal shark habitat within a shallow lagoonal Estuary. PLOS ONE, 13(4), e0195221. https://doi.org/10.1371/journal.pone.0195221
Giaroli, M. L., Byrne, I., Gilby, B. L., Taylor, M., Chargulaf, C. A., & Tibbetts, I. R. (2024). The distribution and significance of stingray feeding pits in Quandamooka (Moreton Bay), Australia. Marine and Freshwater Research, 75(18). https://doi.org/10.1071/mf23247
Heithaus, M. R., Frid, A., Wirsing, A. J., & Worm, B. (2008). Predicting ecological consequences of marine top predator declines. Trends in Ecology & Evolution, 23(4), 202-210. https://doi.org/10.1016/j.tree.2008.01.003
Kolmann, M. A., Huber, D. R., Motta, P. J., & Grubbs, R. D. (2015). Feeding biomechanics of the cownose ray, Rhinoptera bonasus, over ontogeny. Journal of Anatomy, 227(3), 341-351. https://onlinelibrary.wiley.com/doi/full/10.1111/joa.12342
Myers, R. A., Baum, J. K., Shepherd, T. D., Powers, S. P., & Peterson, C. H. (2007). Cascading effects of the loss of APEX predatory sharks from a coastal ocean. Science, 315(5820), 1846-1850. https://doi.org/10.1126/science.1138657
Newell, R. I. (2004). Ecosystem influences of natural and cultivated populations of suspension-feeding bivalve molluscs: A review. 23(1), 51–61. Journal of Shellfish Research, 23(1), 51-61. https://go.gale.com/ps/i.do?id=GALE%7CA118543914
Peterson, C. H., Fodrie, J. F., Summerson, H. C., & Powers, S. P. (2001). Site-specific and density-dependent extinction of prey by schooling rays: generation of a population sink in top-quality habitat for bay scallops. Oecologia, 129, 349-356. https://link.springer.com/article/10.1007/s004420100742
Schwartz, F. J., & Dahlberg, M. D. (1978). Biology and ecology of the Atlantic Stingray, Dasyatis Sabina (Pisces: Dasyatidae) in North Carolina and Georgia. Northeast Gulf Science, 2(1). https://doi.org/10.18785/negs.0201.01
Smith, J. W., & Merriner, J. V. (1985). Food habits and feeding behavior of the Cownose ray, Rhinoptera bonasus, in lower Chesapeake Bay. Estuaries, 8(3), 305. https://doi.org/10.2307/1351491
Smith, J. W., & Merriner, J. V. (1987). Age and growth, movements and distribution of the Cownose ray, Rhinoptera bonasus, in Chesapeake Bay. Estuaries, 10(2), 153. https://doi.org/10.2307/1352180
Snelson, F. F., Williams-Hooper, S. E., & Schmid, T. H. (1988). Reproduction and ecology of the Atlantic Stingray, Dasyatis Sabina, in Florida coastal lagoons. Copeia, 1988(3), 729. https://doi.org/10.2307/1445395
Thrush, S. F., & Dayton, P. K. (2002). Disturbance to marine benthic habitats by trawling and dredging: Implications for marine biodiversity. Annual Review of Ecology and Systematics, 33(1), 449-473. https://doi.org/10.1146/annurev.ecolsys.33.010802.150515
Zu Ermgassen, P. S., Spalding, M. D., Blake, B., Coen, L. D., Dumbauld, B., Geiger, S., Grabowski, J. H., Grizzle, R., Luckenbach, M., McGraw, K., Rodney, W., Ruesink, J. L., Powers, S. P., & Brumbaugh, R. (2012). Historical ecology with real numbers: Past and present extent and biomass of an imperilled estuarine habitat. Proceedings of the Royal Society B: Biological Sciences, 279(1742), 3393-3400. https://doi.org/10.1098/rspb.2012.0313
There are mornings along the edges of the water in Onslow County when the surface looks still enough to trust.
The marsh grass has not yet reached its summer height. What stands there leaves more water exposed between the stems, and without sustained wind, the surface holds its shape. You can see farther into it now than you will in a few weeks, before suspended sediment and constant movement return it to opacity. The water carries less of the season, and because of that, more of what moves beneath it becomes visible—if you are willing to wait long enough to see the difference between movement and reflection.
This is when people begin to notice them again.
Not all at once. Not everywhere. Just a change that does not follow wind or tide. A line that holds where the rest of the surface releases. Something that holds its position in a system that is always adjusting.
An alligator does not arrive in that moment.
It becomes visible.
Alligator emerging from the mud. | Photo credit: Gilbert Grant, iNaturalist
Seasonal Absence Is Not Absence
Through winter, they remain within these same creeks, marsh edges, and quieter channels. What changes is not location, but how they occupy it. As temperatures fall, activity narrows. Movement slows, and the need for it slows with it. Energy is conserved, not spent. And the surface carries fewer signs of what lies beneath it. Individuals hold in deeper water or along softer margins where mud retains heat longer than the surrounding water column, remaining within conditions that allow them to persist without constant movement (Nifong et al., 2014; Rosenblatt & Heithaus, 2011).
The same stretch of water that in spring will hold a visible form can pass through winter without interruption, its stillness mistaken for absence.
But the system does not empty.
It compresses.
The System Wakes in Layers
By early spring, that compression begins to release—not all at once, but in layers that build on each other before they are recognized. Shallow water warms first, taking in solar heat more quickly than deeper channels. Along these edges, fish begin to hold longer. Movements that in winter passed through quickly begin to extend into areas that had remained quiet. Invertebrates return to the sediment surface, and the water column begins to carry more suspended life, even before it becomes visible as turbidity.
Birds respond to this before most other changes are noticed. Their movements tighten. Landings become more frequent, departures more abrupt. What they are tracking is not random. It is the redistribution of energy into places where it can be accessed.
The alligator moves within that shift.
Not as a trigger. Not as something layered on top. But as part of a system reorganizing itself across temperature, light, and movement at the same time.
Great blue heron and alligator are part of an interconnected system. | Photo credit: Audubon North Carolina
Reading What It Is Responding To
When one becomes visible along the edge of a creek or marsh, it is easy to reduce that moment to temperature alone. Warmer water allows for more activity.
But what draws it into that position is more specific than warmth.
It is the arrangement of prey.
Along the margins where water meets land, movement compresses. Fish traveling with the tide encounter shallow gradients that limit how long they can remain. Small mammals moving between marsh and upland must cross exposed edges. Birds landing to feed do so in places where depth and access align for only short intervals.
These are not isolated events. They are recurring patterns shaped by tidal cycles, substrate, and seasonal change.
The alligator positions itself within those patterns.
Its diet reflects that flexibility, spanning invertebrates, fish, birds, reptiles, and mammals depending on size and availability (Nifong, 2016). But the diet alone does not explain its placement. What matters is where energy becomes concentrated, even briefly.
That concentration is not constant. It forms and dissolves with tide, with light, with movement.
And the predator tracks that.
And what appears as a single movement—a fish turning, a bird lifting, something crossing the edge of the marsh—is part of a larger structure that holds only briefly before dissolving again.
The alligator does not respond to the individual movement.
It responds to the pattern that produces it.
Where Freshwater Meets Salt
These are not just places where water mixes.
They are places where movement is forced—and where that movement becomes available to something waiting at the top of it.
There are places along this coastline where those changes concentrate.
At the mouths of creeks, along the edges of the Intracoastal Waterway, and near the shifting bars of New River Inlet, the water does not settle into a single condition. Freshwater moves outward with tide and rainfall, meeting saltwater pressing back in with tidal exchange. The result is not a fixed boundary, but a gradient that shifts continuously—sometimes visible as a faint line, sometimes only detectable in how the surface moves differently from one side to the other.
This is where alligators are most often encountered—because this is where the system compresses into something they can use.
They are not marine animals. They do not possess the specialized salt glands that allow for extended life in high salinity environments. Over time, saltwater carries a physiological cost, requiring a return to freshwater to restore balance (Rosenblatt & Heithaus, 2011; Fujisaki et al., 2014).
But that limitation does not exclude them.
It defines how they move through them.
In these mixing zones, salinity is not constant. It rises and falls with tide, with rainfall, with wind direction. A location that carries higher salinity at one stage may shift toward fresher conditions hours later. What appears to be a boundary is, in practice, a moving field.
Within that field, movement compresses.
Fish traveling with the tide are funneled into narrower pathways. Shallow gradients limit how long they can remain in deeper water. Schools tighten. Individuals encounter edges that restrict escape. The system concentrates energy into space.
The predator does not need to range widely in these conditions.
It needs to hold where movement is forced.
And so it does.
An alligator near the tall grass near Marine Corps Air Station New River | Photo credit: Martin Egnash
At the Edge of the Open Water
There are moments when that pattern extends beyond the mixing zones, into places that appear, at first, outside of where an alligator belongs.
Along the shoreline, in the breaking waves where the ocean meets sand, one will sometimes appear—rising and falling with the swell, holding position just beyond where the water turns over onto the beach. It looks misplaced, as though it has moved beyond the system that defines it.
It has not.
The surf zone is one of the most compressed environments along the coast. Waves reduce depth, disrupt orientation, and concentrate movement into a narrow band where escape is limited. Fish pushed into breaking water lose some ability to maintain direction. Schools fragment. Individuals become briefly exposed in ways that do not occur in deeper, more stable water.
For a predator capable of stillness followed by short bursts of movement, that compression creates opportunity.
But the cost is higher.
Salinity is elevated. The water is in constant motion. There is no stable refuge within immediate reach. Time in this environment cannot be extended indefinitely.
And so it does not.
Movements into higher salinity water tend to be brief—extensions outward, followed by a return to freshwater or lower salinity conditions where balance can be restored (Nifong et al., 2014).
What appears as an anomaly is part of a larger pattern.
The predator crosses the boundary not to remain, but to use it, moving where the system briefly offers more than it costs.
The same forces that shape the marsh edge—compression, constraint, and brief exposure—are recreated here, just for a moment, in a different place.
An alligator rests at the ocean’s edge in North Topsail. | Photo credit: Fox8 Digital Desk
What Its Presence Changes
Most of what that presence changes cannot be seen when it is observed.
Long before any direct interaction occurs, it is already altering how other organisms use space.
Fish moving along the edge do not simply pass through. They adjust their depth, their speed, the amount of time they remain exposed. Birds land with shorter intervals between contact and departure. Mammals approaching the water shift their paths or their timing. These changes are not dramatic in isolation. But they are continuous.
Over time, they accumulate into structure—the kind that determines who feeds, where they feed, and how long they remain.
The influence of a predator at this level extends beyond what it consumes. It shapes behavior across multiple species, redistributing where and how energy moves through the system. The possibility of predation—present even when not observed—alters interactions in ways that regulate access to habitat and resources (Heithaus et al., 2008; Ripple et al., 2014; Estes et al., 2011).
What holds the system in place is not removal alone.
It is pressure.
What is being shaped is not just movement, but access—and access is what determines how energy moves through the system.
More Than Predation
The influence of the alligator does not end with what it hunts, but extends beyond those interactions.
As it moves through shallow systems, it disturbs sediment, creating depressions and pathways that alter how water is retained and how nutrients are redistributed. These small changes in physical structure create conditions that other species use—temporary refuges, feeding areas, and zones where organic material accumulates (Eversole et al., 2018; Subalusky et al., 2009).
In wetland systems, these disturbances have been linked to broader effects, including nutrient cycling and carbon storage, where the presence of large predators contributes to the retention of organic material within the system rather than its export (Murray et al., 2025; Atwood et al., 2015).
These processes do not occur in isolation.
They intersect with the same patterns of movement, feeding, and behavior that define the system at larger scales.
Seeing the Surface, Reading the System
When one becomes visible along the surface, it is easy to treat the moment as singular.
A sighting. An encounter. Something separate from everything around it.
But that form at the surface is supported by layers extending beyond what can be seen.
It reflects water temperatures crossing into ranges that support sustained activity. It reflects prey moving into positions where access becomes possible. It reflects a system where behavior is still shaped by the presence of something at the top.
The alligator is not an interruption to that system.
It is an expression of it.
What Becomes Visible
Seeing one does not indicate that something has entered the water.
It indicates that enough beneath the surface is functioning to hold it.
Not in a static sense. Not as balance in the way it is often described. But as a set of interactions that remain connected—movement, response, pressure—each shaping the others even when they are not directly observed.
What becomes visible at the surface is only a fraction of that structure.
But it is enough to know that the rest is still in place.
An alligator in Onslow County sits at the edge of the saltmarsh. |Photo credit: Gilbert Grant, iNaturalist
When That Pressure Is Reduced
If that pressure is reduced, the system does not leave an obvious gap.
It shifts.
Movements that were once constrained begin to extend. Species that passed quickly through exposed areas begin to remain longer. Edges that functioned as transition zones become used differently—not because the physical environment has changed, but because the conditions that shaped behavior within it have relaxed.
Mid-level predators expand their activity under these conditions, increasing their access to prey and space when not constrained from above (Nifong et al., 2013).
The change is subtle.
It appears in how long something stays. In how often it returns. In where it lingers. In how quietly the structure of behavior begins to loosen.
The food web and trophic cascade of the American alligator in the Florida Everglades.
A System Written Into Temperature
There is another layer to this that does not show itself at the surface.
The structure of that presence is set years earlier, in a place that can be overlooked when standing at the water’s edge. Along the margins of marsh and wetland, slightly above the reach of regular water movement, nests are built from vegetation and sediment, forming mounds that hold heat as they decompose.
Within those mounds, temperature determines something that will not be visible for much later.
Sex is not fixed at fertilization. It emerges during incubation, shaped by the thermal conditions held within the nest. A difference of only a few degrees is enough to shift the outcome, producing more males or more females depending on where within that range the nest remains (Lang & Andrews, 1994; Janzen, 1994).
Under variable conditions—differences in shading, rainfall, timing, and placement—those outcomes are distributed across the landscape. Some nests produce more females, others more males. That variability holds the population in a form that can sustain itself over time.
When conditions become more consistent, that variation narrows.
Warmer nights hold heat longer within the nest. Seasonal transitions extend. The range of outcomes compresses. What was once distributed begins to align.
And that alignment carries forward into the structure of the population—into how individuals occupy space, into how pressure is applied across the system, into what will eventually be visible at the surface.
Alligator eggs hatch after 65 days of incubation in the fall. The babies will chirp to alert their mom, who then digs out the nest while the babies use their egg tooth to hatch from their eggs. Their mom will then safely carry them to the water.
Where the Next Generation Is Set
The placement of those nests depends on something even more constrained.
A narrow band of land that remains above water just long enough to hold them.
That band is not fixed.
It shifts with tide, with rainfall, with the gradual reworking of shoreline that occurs across seasons and years. With rising sea levels, water reaches farther into areas that once remained above it. Flooding becomes more frequent, not always through singular events, but through repeated intrusions that saturate and destabilize what had previously held (Joanen & McNease, 1989; Sweet et al., 2022).
Human alteration compresses this space further.
Hardened shorelines, dredging, and development reduce the gradual transition between land and water. Where there was once a slope capable of holding multiple elevations, there becomes a defined edge. That edge does not provide the same range of conditions required for successful nesting.
The number of suitable sites decreases.
More importantly, the variability between them narrows.
And with that, the system loses one of the mechanisms that allowed it to absorb change.
Alligator on her nest that can hold up to 60 eggs. | Photo credit: National Park Service (NPS)
What Its Presence Means
When an alligator becomes visible along the surface, it reflects conditions that have aligned across multiple layers.
Temperature has reached a range that supports activity. Prey has moved into positions where access becomes possible. Behavioral pressure remains in place across the system. Reproduction has held across enough years, in enough suitable places, to sustain what is now present.
What is seen at the surface is not separate from them.
It is supported by them.
Seeing one does not signal that something has entered the water.
It signals that enough of what lies beneath it—movement, pressure, response, and continuity—remains intact.
And that—even when most of it is not visible—the system is still holding together.
And that is what becomes visible—just long enough to be seen, before the system closes back over it again.
The system does not end at the water’s edge.
Epilogue: Chicken Nugget
We came across him along the New River, near the courthouse in Jacksonville.
We were there to clear what had been left behind—fishing line caught along the walkways, hooks, and the overflow from a trash can that had spilled out onto the edge. Fast food containers, grocery store chicken trays, scattered along the bank. The signs were clear enough. People had been there for a while—crabbing, fishing, eating, leaving what remained.
He was directly below us.
Small enough to miss at first. Still enough to blend into the water until you stopped looking for movement and started noticing what held its position.
A juvenile alligator, watching.
He stayed there while we worked, then slipped beneath the surface and crossed the small bay. On the opposite side, someone tossed a piece of food into the water. He surfaced almost immediately, took it, and remained.
Waiting.
I came back later and stayed longer.
The pattern repeated. He would disappear until footsteps approached, then return to the same place along the edge. Holding position. Watching. Waiting for something to fall.
No fishermen or crabbers passed through while I was there, but the behavior was consistent with what happens when food becomes predictable. Bait, catch, scraps—anything that can be taken without the cost of searching or pursuing.
Energy, without effort.
It is easy to see something like that and respond to what it looks like in that moment. A small animal. Still. Attentive. Something that feels close enough to interact with.
But what is being shaped there is not just a single interaction.
It is behavior.
A shift away from the conditions that formed it—toward something more efficient, more immediate, and less stable over time. The system that once required movement, patience, and response begins to narrow into expectation.
And expectation changes how an animal uses space.
What happens when that animal is no longer small is not a separate question.
It is the continuation of the same pattern.
Alligators do not forget where food has been easy to obtain. They return to it. They hold in those places. They begin to associate presence—human presence—with opportunity.
What begins as something that feels harmless becomes something that alters how the system functions around it.
Not just for the animal, but for everything that responds to it.
There are instincts at work here that were shaped long before any walkway, any dock, any place where food might be dropped from above. Those instincts are not just about survival in isolation. They are part of how pressure is applied, how movement is shaped, how the system holds.
When those instincts are replaced with something easier, the effect does not remain contained.
It carries outward.
He stayed there while I watched. Returning to the same place. Holding the same position. Waiting for something to fall.
There is a kind of kindness in wanting to give something to an animal like that.
But there is another kind in leaving it as it is.
Not interrupting the conditions that shape it. Not narrowing what it has learned to expect. Not replacing a system built on movement and response with one built on waiting.
Let it remember the water as it is.
And you, only as something that passed through it.
We affectionately named this juvenile alligator in the New River in Jacksonville, NC “Chicken Nugget” for all of the chicken nugget boxes left behind on the walkway from an overflowing trash can. | Photo credit: A. Mitchell
References
Atwood, T. B., Connolly, R. M., Ritchie, E. G., Lovelock, C. E., Heithaus, M. R., Hays, G. C., Fourqurean, J. W., & Macreadie, P. I. (2015). Predators help protect carbon stocks in blue carbon ecosystems. Nature Climate Change, 5(12), 1038-1045. https://doi.org/10.1038/nclimate2763
Estes, J. A., Terbough, J., Brashares, J. S., Power, M. E., Berger, J., Bond, W. J., Carpenter, S. R., Essington, T. E., Holt, R. D., & Wardle, D. A. (2011). Trophic Downgrading of Planet Earth. Science, 333(604), 301-306. https://www.science.org/doi/abs/10.1126/science.1205106
Fujisaki, I., Hart, K. M., Mazzotti, F. J., Cherkiss, M. S., Sartain, A. R., Jeffery, B. M., Beauchamp, J. S., & Denton, M. (2014). Home range and movements of American alligators (Alligator mississippiensis) in an Estuary habitat. Animal Biotelemetry, 2(1), 8. https://doi.org/10.1186/2050-3385-2-8
Heithaus, M. R., Frid, A., Wirsing, A. J., & Worm, B. (2008). Predicting ecological consequences of marine top predator declines. Trends in Ecology & Evolution, 23(4), 202-210. https://doi.org/10.1016/j.tree.2008.01.003
Janzen, F. J. (1994). Climate change and temperature-dependent sex determination in reptiles. PNAS, 91(16), 7487-7490. https://doi.org/10.1073/pnas.91.16.7487
Joanen, T., & McNease, L. L. (1989). Ecology and physiology of nesting and early development of the American alligator. American Zoologist, 29(3), 987-998. https://doi.org/10.1093/icb/29.3.987
Lang, J. W., & Andrews, H. V. (1994). Temperature‐dependent sex determination in crocodilians. Journal of Experimental Zoology, 270(1), 28-44. https://doi.org/10.1002/jez.1402700105
Nifong, J. C. (2016). Living on the edge: Trophic ecology of alligator mississippiensis (American alligator) with access to a shallow estuarine impoundment. Bulletin of the Florida Museum of Natural History, 54(2), 13-49. https://doi.org/10.58782/flmnh.xkdw7119
Nifong, J. C., Nifong, R. L., Silliman, B. R., Lowers, R. H., Guillette, L. J., Ferguson, J. M., Welsh, M., Abernathy, K., & Marshall, G. (2014). Animal-borne imaging reveals novel insights into the foraging behaviors and Diel activity of a large-bodied APEX predator, the American alligator (Alligator mississippiensis). PLoS ONE, 9(1), e83953. https://doi.org/10.1371/journal.pone.0083953
Nifong, J. C., & Silliman, B. R. (2013). Impacts of a large-bodied, APEX predator (Alligator mississippiensis Daudin 1801) on salt marsh food webs. Journal of Experimental Marine Biology and Ecology, 440, 185-191. https://doi.org/10.1016/j.jembe.2013.01.002
Ripple, W. J., Estes, J. A., Beschta, R. L., Wilmers, C. C., Ritchie, E. G., Hebblewhite, M., Berger, J., Elmhagen, B., Letnic, M., Nelson, M. P., Schmitz, O. J., Smith, D. W., Wallach, A. D., & Wirsing, A. J. (2014). Status and ecological effects of the world’s largest carnivores. Science, 343(6167). https://doi.org/10.1126/science.1241484
Rosenblatt, A. E., & Heithaus, M. R. (2011). Does variation in movement tactics and trophic interactions among American alligators create habitat linkages? Journal of Animal Ecology, 80(4), 786-798. https://doi.org/10.1111/j.1365-2656.2011.01830.x
Sweet, W. V., Hamlington, B. D., Kopp, R. E., Weaver, C. P., Barnard, P. L., Bekaert, D., Brooks, W., Craghan, M., Dusek, G., Frederickse, T., Garner, G., Genz, A. S., Krasting, J. P., Larour, E., Marcy, D., Marra, J. J., Obeysekera, J., Osler, M., Pendleton, M., … Zuzak, C. (2022). Global and regional sea level rise scenarios for the United States: Updated mean projections and extreme water level probabilities along U.S. coastlines (Technical Report NOS 01). National Oceanic and Atmospheric Administration, National Ocean Service. https://earth.gov/sealevel/us/internal_resources/756/noaa-nos-techrpt01-global-regional-SLR-scenarios-US.pdf
There are stretches of shoreline in Onslow County where the water looks simple.
A low wind flattens the surface just beyond the breakers. The sand underfoot is firm, packed by a falling tide. Small schools of baitfish turn in unison at the edge of visibility, their bodies catching light and then disappearing again as if nothing had moved at all.
From here, fish seem predictable. They swim. They are streamlined. They slip through water in ways that feel consistent, almost mechanical.
A school of juvenile fish swim in the Surf City sound. | Photo credit: A. Mitchell
But that impression doesn’t hold for long.
A few steps into the surf, something crunches beneath your heel—a shell, or what remains of one.. Offshore, a shape drifts that doesn’t seem built for movement at all. In the shallows, something settles to the bottom and then, impossibly, walks.
The closer you look, the more the pattern breaks apart. Along this stretch of coast—from the swash zone to the deeper water of Onslow Bay—some fish are not built like fish are “supposed” to be.
And once you notice them, the rules start to feel less like rules at all.
Teeth built for stone: Sheepshead
The rule broken: fish are supposed to have simple teeth
On calm mornings near New River Inlet, when the tide is just beginning to push in, the water around pilings and rock edges clears enough to see movement below the surface. Dark vertical bands appear and disappear as fish turn sideways to feed, their bodies angled tightly against pilings and rock.
If you get a close look—often only when one is caught—you notice the teeth.
Flat. Squared. Set in rows that look more like something borrowed from a mammal than a fish.
Sheepshead fish have mammal-like teeth used for scraping and crushing hard shells and barnacles. | Photo credit: Jeannette’s PIer
The sheepshead (Archosargus probatocephalus) feeds primarily on hard-shelled organisms—barnacles, oysters, mussels, and crabs attached to pilings, jetties, and natural hardbottom (Sedberry, 1987). These prey items are abundant in estuarine and nearshore environments where salinity fluctuates and structure concentrates life.
Instead of pointed, uniform teeth, sheepshead possess incisiform front teeth for scraping and strong molariform teeth set further back for crushing (Deang et al., 2018; Hernandez & Motta, 1997). Bite force measurements and stomach content analyses show they are capable of breaking calcareous shells that would resist most coastal fishes (Hernandez & Motta, 1997).
They are most active in waters typically ranging from 60–80°F (15–27°C), often within just a few feet of structure in depths from less than a meter to roughly 10 meters (Sedberry, 1987).
Fish are often imagined as generalized swimmers feeding on soft prey. But along the Onslow coast, hard surfaces—oyster beds, submerged debris, pilings—create entire microhabitats built on calcium carbonate (Grabowski & Peterson, 2007).
Sheepshead are not exceptions to the system; they are shaped by it. Their teeth are a direct response to a landscape where food remains locked inside a shell.
Most fish don’t have teeth like this because most environments don’t require it. Here, where geology and biology meet in layers of shell and structure, the rule changes.
The fish that walks: Bluespotted and Northern searobin
The rule broken: fish move by swimming
On a falling tide along the edges of Topsail Island, the water pulls thin over the sand flats. What remains is a shifting surface—ripples, shadows, and the occasional sudden burst of motion.
Then something moves without swimming.
It doesn’t dart or glide. It advances in short, deliberate steps, stopping and starting again, as if testing the ground before each movement.
For a moment, it looks wrong—like something moving through air instead of water.
The bluespotted searobin (Prionotus roseus) and the Northern searobin (Prionotus carolinus) do not rely on their fins for propulsion in the way most fish do. Instead, three detached rays from each pectoral fin extend downward, contacting the bottom and supporting the body as it moves. These rays function both as supports and as sensory structures, probing the sediment and detecting chemical cues—effectively allowing the fish to “taste” the seafloor as it moves (Bardach & Case, 1965).
Across these shallow flats, often just inches to a few feet deep, the water warms into the upper 60s and 70s as the tide recedes. Prey is rarely exposed. Worms, small crustaceans, and buried mollusks remain hidden beneath the surface. Vision alone is not enough here. The searobin moves slowly, stepping and pausing, tracing the bottom until something beneath the sand gives itself away.
Movement in water is usually about efficiency—minimizing drag, maximizing speed.
But the seafloor is a different environment entirely.
Here, visibility narrows, prey disappears beneath the surface, and swimming can carry you past what you’re trying to find. Walking—slow, deliberate, sensory-driven—becomes the better strategy.
Most fish don’t have “legs” because most fish don’t live where walking is more useful than swimming. Along the shallow bottoms of Onslow waters, this rule no longer applies.
The fish that swells: Northern puffer
The rule broken: fish don’t change shape
In late summer, when the water just beyond the breakers settles into the upper 70s, small shapes begin to move just offshore—slow, almost indifferent to the motion around them.
One drifts closer than expected, rounded in a way that doesn’t quite match the others. It hovers, turning slightly, its movement controlled but unhurried.
Then, without warning, the body changes.
It expands outward, the outline swelling until the fish no longer resembles something built to move through water at all.
The Northern puffer (Sphoeroides maculatus) does this by rapidly drawing water into a highly elastic stomach, a process that allows the body to expand far beyond its resting shape (Brainerd, 1994). Without rigid skeletal constraints like ribs or pelvic bones, that expansion can happen quickly, transforming the fish into something difficult for a predator to grasp or swallow.
A Northern pufferfish skeleton is made up of spiny modified scales (not bones) that expand like a balloon when threatended. | Photo credit: The Fossil Forum
In these nearshore waters—where predators move quickly and encounters happen at close range—there is little time to outrun what’s coming. Most fish rely on speed to escape. This one changes shape instead.
Speed isn’t part of the solution here.
The fish that locks itself in place: Gray triggerfish
The rule broken: fish don’t anchor themselves
Farther offshore, where the bottom begins to break into scattered hardbottom and reef patches, movement slows in a different way.
Shapes hold just above the structure, adjusting position in small increments, never straying far from the surface below them.
When disturbed, they don’t flee into open water.
They turn downward.
The gray triggerfish (Balistes capriscus) moves into crevices and tight spaces within the structure, where a set of dorsal spines can be raised and locked into place. The first spine lifts, and a smaller second spine holds it there—an arrangement that gives the fish its name and allows it to anchor itself firmly in place (Tyler, 1980; Lobel, 1980).
Its body is built for this kind of movement: deep and laterally compressed, with tough, abrasive skin and strong incisor-like teeth capable of breaking into hard-shelled prey (Tyler, 1980; Lobel, 1980). These are not features meant for speed. They are features meant for contact—pressing into structure, resisting removal, holding position when movement would fail.
In waters often 50–120 feet deep off Onslow County, where reefs and wrecks break the seafloor into pockets and edges, escape doesn’t always mean distance (Bellwood et al., 2004).
Sometimes it means holding ground.
Most fish survive by staying in motion.
This one survives by becoming fixed in place, turning the structure around it into part of its defense.
Light written into skin: Atlantic midshipman
The rule broken: fish don’t carry light in their skin
On warm summer nights near quiet stretches of marsh and inlet edges, the water sometimes carries sound before anything else. A low, continuous hum. It’s easy to miss unless you stop moving.
The Atlantic midshipman (Porichthys plectrodon) produces that sound through specialized sonic muscles vibrating against the swim bladder, creating a sustained hum that can carry through shallow coastal water (Sisneros, 2009; Bass & McKibben, 2003).
If you listen carefully during a quiet evening, the sound of a male midshipman trying to court a female might be heard. | Audio credit: SanctoSound – Integrated Ocean Observing System (IOOS)
Along the sides of the body and across the head are rows of small organs—photophores—set into the skin, giving the fish its name and marking it as something unusual among coastal species found in these waters (Schwartz, 2013). When seen out of the water, those rows catch the light in a very particular way—small, round points that flash gold in direct sunlight, spaced with a regularity that makes them look almost set into the surface, like buttons fixed into the skin.
The Atlantic midshipman has photophores that dazzle when out of the water, and used in seeing in darkened burrows and structures in limited light. | Photo credit: North American Native Fishes Association
Midshipman inhabit shallow coastal environments, often in burrows or beneath structure along muddy or sandy bottoms, typically in depths less than 20 meters.
Light in fish is often associated with deeper water, where darkness is constant and illumination becomes necessary (Haddock et al., 2010). But along the Onslow coast, those conditions can exist in smaller, shifting pockets. Light narrows quickly with depth, suspended sediment moves with the tide, and visibility can collapse even in water shallow enough to stand in.
Not all fish in these waters experience the bottom the same way. A flounder rests exposed on the sand, relying on camouflage and stillness. The midshipman, by contrast, spends much of its time within burrows, beneath structure, or pressed close to the substrate, where light is already limited and often disappears entirely.
In those spaces, the rules of visibility begin to resemble something closer to deeper water, even though the surface is only a few feet above.
The presence of photophores here does not follow the pattern most people expect.
Not all light comes from above.
The deep blade: Long-snouted lancetfish
The rule broken: fish are dense, muscular swimmers
From the beach, the horizon feels like a boundary—beyond the sandbars, beyond the nearshore currents—about two miles out, where the surface lifts just enough to hide what comes after. But beyond that line, the water doesn’t simply continue. It changes.
Depth increases quickly. Layers begin to form. Light fades long before the bottom is reached.
And in those deeper waters off Onslow Bay, some fish are not built to chase anything at all.
The long-nosed lancetfish lives in the middle depths of the ocean where body density is less desirable for a drifting fish. | Photo credit: ML – some rights reserved (CC BY-NC)
The long-snouted lancetfish (Alepisaurus ferox) lives in the midwater column, often hundreds of meters below the surface. Its body is long and thin, almost blade-like, with muscle reduced and tissue that is less dense than most active predators, appearing almost soft in the water (Drazen & Seibel, 2007).
It does not move with the steady, powered swimming most fish rely on. Instead, it drifts, adjusting position and taking prey as it comes within reach. Stomach analyses show a wide range of prey—fish, squid, and even other lancetfish—suggesting opportunism rather than pursuit (Kubota & Uyeno, 1970).
In these deeper layers, energy becomes harder to acquire and more costly to use.
Building and maintaining dense muscle comes at a cost. Chasing prey demands more of it (Sutton, 2013).
Here, that balance shifts.
The lancetfish represents a different solution—one that reduces the cost of movement and relies instead on encounter.
Most fish are built to swim.
This one is built to wait.
The armored survivor: Atlantic sturgeon
The rule broken: fish are supposed to have scales
In cooler months, when water temperatures drop into the 50s and 60s, large shapes move along the bottom of estuaries and nearshore waters.
They do not flash or turn sharply. They move steadily, close to the sediment.
At times, that movement reaches the surface. A back breaks through, arcing briefly before slipping under again, the shape unfamiliar enough that it doesn’t immediately read as a fish.
The Atlantic sturgeon (Acipenser oxyrinchus oxyrinchus) retains an older form—rows of bony scutes instead of the flexible scales seen in most fishes (Bemis et al., 1997). Along the underside, a protrusible mouth extends downward, drawing in prey from the bottom through suction rather than pursuit (ASSRT, 2007; Bemis et al., 1997).
An anadromous fish, they move between river systems and coastal waters, passing through estuaries and along the nearshore edge, often in depths ranging from shallow channels to over 100 feet offshore (Dunton et al., 2015; ASSRT, 2007).
This design is not new. It has persisted for tens of millions of years, carried forward through changing coastlines, shifting sea levels, and the rise of entirely different groups of fishes (Bemis et al., 1997).
Atlantic sturgeon have a bony structure that has remained relatively unchanged for millions of years. | Photo credits: mdadswell – some rights reserved (CC BY-NC) (left); Steven McGrath – some rights reserved (CC BY-NC-ND) (right)
It works because the conditions it responds to have never fully disappeared.
Along the bottom, prey remains buried. Sediment still shifts with current and tide. Feeding still depends on contact more than speed. Armor still protects a body that cannot easily maneuver out of danger.
For a long time, these fish seemed to fade from local waters. In Onslow County, encounters became rare enough to feel like absence. But populations have persisted elsewhere, and in nearby systems like the Cape Fear River, they are being observed again with increasing frequency—moving through channels, returning to spawning grounds, reappearing in places where they had not been seen in years (Dunton et al., 2015; ASSRT, 2007).
Their range has shifted before. It may be shifting again.
What remains constant is the need for connection—between river and ocean, between spawning grounds and feeding habitat.
This fish does not depend on a single place. It depends on the continuity between them.
Not all designs are meant to change. Some persist because the system they belong to still exists.
The drifting giant: Ocean sunfish
The rule broken: fish are supposed to be shaped for swimming
Occasionally, especially in warmer months when currents shift, something appears offshore that barely seems to move at all.
A large, flattened body. A fin breaking the surface. Then another, held there longer than expected.
It drifts more than it swims.
At times, it lingers there, tilted at the surface, absorbing the sun before slipping back beneath the water.
The ocean sunfish (Mola mola) is one of the heaviest bony fish, reaching weights over 1,000 kg. Its body is truncated, lacking a true caudal fin, and propulsion is achieved through synchronized movements of dorsal and anal fins (Pope et al., 2010; Watanabe et al., 2009).
After diving into colder, deeper water, sunfish often return to the surface, where this slow, drifting posture allows their body temperature to rise again (Watanabe et al., 2009). Prolonged time at the surface can leave the skin visibly altered—shifting from darker grey to lighter tones, sometimes appearing pale or pinkened under sustained exposure.
Sunfish often inhabit offshore waters but can approach nearshore areas following currents and prey, particularly gelatinous organisms like jellyfish (Cartamil & Lowe, 2004).
By most expectations, this body plan shouldn’t work.
But it does—because efficiency, here, takes a different form. It is about buoyancy, drift, and feeding on abundant, slow-moving prey.
In a system where jellyfish blooms are seasonal and sometimes dense, a fish shaped like this becomes not an anomaly, but a specialist.
Answers to a layered environment
From the shoreline, the water still looks simple.
Small waves rise and fall. Baitfish turn and vanish. The surface holds its shape.
But beneath that surface, the rules have already begun to shift.
Fish move through these waters in ways that don’t match what we expect—crushing shell, stepping across the bottom, changing shape, holding themselves in place, carrying structures that catch light, drifting where others would swim, or moving through forms shaped long before this coastline took its present shape.
What appears, from the beach, to be a single environment is something else entirely. It is layered—sand, structure, depth, temperature, light—each one asking something different of the animals that live within it.
And the fish that seem unusual are not exceptions.
They are answers.
A layered system, at New River Inlet, seen from the surface. | Photo credit: A. Mitchell
Bardach, J. E., & Case, J. (1965). Sensory capabilities of the modified fins of squirrel hake (Urophycis chuss) and Searobins (Prionotus carolinus and P. evolans). Copeia, 1965(2), 194. https://doi.org/10.2307/1440724
Bellwood, D. R., Hughes, T. P., Folke, C., & Nyström, M. (2004). Confronting the coral reef crisis. Nature, 429(6994), 827-833. https://doi.org/10.1038/nature02691
Bemis, W. E., Findeis, E. K., & Grande, L. (1997). An overview of Acipenseriformes. Developments in Environmental Biology of Fishes, 48, 25-71. https://doi.org/10.1007/0-306-46854-9_4
Brainerd, E. L. (1994). Pufferfish inflation: Functional morphology of postcranial structures in Diodon holocanthus (Tetraodontiformes). Journal of Morphology, 220(3), 243-261. https://doi.org/10.1002/jmor.1052200304
Cartamil, D., & Lowe, C. (2004). Diel movement patterns of ocean sunfish mola mola off Southern California. Marine Ecology Progress Series, 266, 245-253. https://doi.org/10.3354/meps266245
Deang, J., Persons, A., Oppedal, A., Rhee, H., Moser, R., & Horstemeyer, M. (2018). Structure, property, and function of sheepshead (Archosargus probatocephalus) teeth. Archives of Oral Biology, 89, 1-8. https://doi.org/10.1016/j.archoralbio.2018.01.013
Drazen, J. C., & Seibel, B. A. (2007). Depth‐related trends in metabolism of benthic and benthopelagic deep‐sea fishes. Limnology and Oceanography, 52(5), 2306-2316. https://doi.org/10.4319/lo.2007.52.5.2306
Dunton, K. J., Jordaan, A., Conover, D. O., McKown, K. A., Bonacci, L. A., & Frisk, M. G. (2015). Marine distribution and habitat use of Atlantic sturgeon in New York lead to fisheries interactions and Bycatch. Marine and Coastal Fisheries, 7(1), 18-32. https://doi.org/10.1080/19425120.2014.986348
Fernandez, L. P., & Motta, P. J. (1997). Trophic consequences of differential performance: Ontogeny of oral jaw‐crushing performance in the sheepshead, Archosargus probatocephalus (Teleostei, sparidae). Journal of Zoology, 243(4), 737-756. https://doi.org/10.1111/j.1469-7998.1997.tb01973.x
Grabowski, J. H., & Peterson, C. H. (2007). Restoring oyster reefs to recover ecosystem services. Theoretical Ecology Series, 4, 281-298. https://doi.org/10.1016/s1875-306x(07)80017-7
McIver, E. L., Marchaterre, M. A., Rice, A. N., & Bass, A. H. (2014). Novel underwater soundscape: Acoustic repertoire of plainfin midshipman fish. Journal of Experimental Biology. https://doi.org/10.1242/jeb.102772
Mensinger, A. F., & Case, J. F. (1990). Luminescent properties of deep sea fish. Journal of Experimental Marine Biology and Ecology, 144(1), 1-15. https://doi.org/10.1016/0022-0981(90)90015-5
Petersen, J. C., & Ramsay, J. B. (2020). Walking on chains: The morphology and mechanics behind the fin ray derived limbs of sea-robins. Journal of Experimental Biology. https://doi.org/10.1242/jeb.227140
Peterson, D. L., Bain, M. B., & Haley, N. (2000). Evidence of declining recruitment of Atlantic sturgeon in the Hudson River. North American Journal of Fisheries Management, 20(1), 231-238. https://doi.org/10.1577/1548-8675(2000)020<0231:eodroa>2.0.co;2
Pope, E. C., Hays, G. C., Thys, T. M., Doyle, T. K., Sims, D. W., Queiroz, N., Hobson, V. J., Kubicek, L., & Houghton, J. D. (2010). The biology and ecology of the ocean sunfish mola mola: A review of current knowledge and future research perspectives. Reviews in Fish Biology and Fisheries, 20(4), 471-487. https://doi.org/10.1007/s11160-009-9155-9
Schwartz, F. J. (2013). Atlantic midshipman, Porichthys plectrodon, in North Carolina. Journal of the North Carolina Academy of Science, 129(3), 111-114. https://doi.org/10.7572/2167-5880-129.3.111
Sedberry, G. R. (1987). Feeding habits of Sheepshead, Archosargus probatocephalus, in offshore reef habitats of the southeastern continental shelf. Northeast Gulf Science, 9(1). https://doi.org/10.18785/negs.0901.03
Sisneros, J. A. (2009). Adaptive hearing in the vocal plainfin midshipman fish: Getting in tune for the breeding season and implications for acoustic communication. Integrative Zoology, 4(1), 33-42. https://doi.org/10.1111/j.1749-4877.2008.00133.x
Snelgrove, P. V. (1999). Getting to the bottom of marine biodiversity: Sedimentary habitats. BioScience, 49(2), 129. https://doi.org/10.2307/1313538
Sutton, T. T. (2013). Vertical ecology of the pelagic ocean: Classical patterns and new perspectives. Journal of Fish Biology, 83(6), 1508-1527. https://doi.org/10.1111/jfb.12263
Tyler, J. C. (1980). Osteology, phylogeny, and higher classification of the fishes of the order plectognathi (Tetraodontiformes) (434). U.S. Dept. of Commerce, National Oceanic and Atmospheric Administration, National Marine Fisheries Service. https://10.5962/bhl.title.63022
Each March 14, mathematicians celebrate π — the constant that links the circumference of a circle to its diameter. But Pi Day in nature appears everywhere along the coast: in boundaries that curve back upon itself, in ripples spreading across still water, in the rounded mouth of a burrow, in the arcs traced by a turning tide. Along the coast, these circles and spirals reveal patterns in nature that emerge so often they begin to feel less like abstract mathematics and more like a language written into sand and water. The shoreline is not calculating anything deliberately, yet the same relationships appear again and again as tides move sediment, organisms grow, and currents redistribute energy. What looks at first like scattered shapes — a curved creek channel, a ring of crab pellets, the fivefold symmetry of a sea star — gradually reveals itself as part of a larger pattern. The coast is full of geometry, briefly visible each time the water recedes.
The Creek Writes in Curves
A tidal creek bends around the marsh edge behind Surf City, where vegetation and sediment redirect the flow of draining water. These shifting boundaries gradually guide channels into widening curves that reappear with each tide. | Photo credit: A. Mitchell
At the creek mouths behind Topsail Island, the marsh edge redraws itself each time the tide drains away. Water retreats through narrow runnels that refuse straight lines, bending around grass hummocks and soft ridges, leaving a fan of nested arcs etched into exposed mud. The channels widen as velocity drops, sediment settling in fractions that record the rate of energy loss, so the surface becomes a temporary map of fluid negotiation.
These curves appear wherever moving water gradually redistributes energy rather than releasing it abruptly. In tidal landscapes, vegetation and sediment interact with flow in feedback loops that reshape channels over time, producing curved drainage networks whose geometry reflects both plant resistance and water momentum (Kirwan & Murray, 2007; Temmerman et al., 2007; Murray & Paola, 1994). Across river basins and tidal creeks alike, these evolving paths often approach widening spiral-like patterns as flow repeatedly adjusts to the boundaries around it (Rodriguez-Iturbe & Rinaldo, 1998).
Foam left behind by the falling tide sometimes dries into thin white filaments that trace these curves for a few quiet minutes before collapsing, a temporary record of motion fixed long enough to be read.
The creek does not preserve a single spiral. Each tide erases and redraws the same proportional tendency. The form emerges not from design but from the repeated redistribution of energy through water and sediment.
Geometry in the Grass
Dense stands of Spartina alterniflora divide space through repeating stem spacing. This structure slows water movement and traps suspended sediment, linking plant growth to the gradual elevation of the marsh surface. | Photo credit: A. Mitchell
Along the marsh margin, stems of Spartina alterniflora divide space through incremental adjustment. Leaves diverge from one another at angles that reduce overlap, distributing light capture through the canopy in repeating offsets that resemble packing patterns seen throughout plant growth.
Experiments in plant development show that when new structures arise under simple inhibitory fields, spiral-like arrangements often emerge as stable growth solutions (Douady & Couder, 1996). These patterns are widely recognized in plant morphology, where spacing between leaves or stems tends to distribute light and nutrients efficiently through the canopy (Niklas, 1997).
In salt marshes, this spacing carries ecological consequences beyond plant structure. Vegetation alters local water flow, slowing currents and promoting the deposition of suspended sediments that gradually elevate the marsh surface (Bouma et al., 2009; Fagherazzi et al., 2013; Leonard & Luther, 1995).
Mud crab burrows often appear in clusters whose spacing echoes the density of surrounding vegetation, each opening maintaining just enough distance to avoid collapse into the next.
Spiral shell growth of the periwinkle snail follows a repeating geometric expansion, allowing the animal to grow while maintaining the same overall shape. | Photo credit: A. Mitchell
Marsh periwinkles climb these stems in staggered lines that mirror the spacing of the leaves, their positions shifting with the tide yet repeatedly settling into the same angular arrangement.
Across the marsh platform, geometry quietly mediates the relationship between plant growth and landscape formation.
Spheres at the Mouth of a Burrow
A mud crab burrow at the edge of marsh vegetation marks the boundary between sand, grass, and moving water where patterns of spacing emerge. | Photo credit: A. Mitchell
Along the upper edge of the beach where grasses begin to anchor the sand, small clusters of rounded pellets often surround the entrances to crab burrows. At first glance they resemble scattered grains or fragments of dry sediment, but kneeling close reveals a more deliberate pattern.
Each pellet forms as damp sand excavated from underground tunnels passes through the crab’s mouthparts before being pushed back to the surface (Lucrezi et al., 2009). As the grains are rolled and compressed together, they settle into rounded shapes before drying in the coastal wind.
Among all possible forms loose material might take, the sphere encloses volume while minimizing surface area — a principle known as the isoperimetric property. When damp sand is compacted from many directions, the grains naturally settle toward this configuration.
The crab does not deliberately engineer spheres; the physics of granular material does the work. Similar rounding appears wherever particles compress together, from bubbles forming in foam to droplets condensing in clouds.
Around the burrow entrance, the pellets accumulate in loose arcs or clustered rings marking the repeated path of excavation. Studies of mud and ghost crab burrowing show that these excavated pellets form characteristic surface patterns around burrow openings as crabs repeatedly transport sediment from their tunnels (Lim & Diong, 2003; Chan et al., 2006).
Within hours the pellets dry and crumble back into ordinary sand. By the next tide the pattern may vanish entirely, erased by waves or shifting grains. Yet while they last, these small spheres record the intersection of animal behavior, sediment physics, and geometry.
Fivefold Bodies in the Wrack
Sand dollars show pentaradial symmetry — a five-part body plan shared by many echinoderms. The familiar white “sand dollar” is the skeleton left behind after the animal dies. Living sand dollars are gray or brown and covered in tiny moving spines that allow them to feed and move through the sand. In North Carolina, collecting live sand dollars is illegal; only empty tests found on the beach may be taken.| Image credit: Suzanne Campbell-O’Rahilly
Along the wrack line, sea stars rest without a preferred direction, their five arms distributing contact evenly across wet sand. Pentaradial symmetry divides the body into five equal sectors, stabilizing locomotion and feeding while allowing regeneration to proceed without disrupting balance (Beadle, 1989).
A broken sea star missing an arm still preserves the angle of the remaining four. The body reorganizes around absence without abandoning its underlying symmetry.
Sand dollars flatten this same geometry into a disk etched with five petal-like openings across the shell surface. These structures guide water across respiratory tissues while reinforcing the skeleton against bending forces generated by waves and sediment movement (Ellers & Telford, 1992; Mooi & David, 1998; Telford, 1981).
In shallow swash zones, freshly uncovered sand dollars often rotate edgewise until resistance equalizes, their circular outlines turning slowly with each pulse of water.
The etched flower is neither ornament nor accident. It records the intersection of circulation and structural strength — a geometry recalculated as abrasion reshapes the shell and burial depth shifts with each surge.
Across many biological systems, similar proportional relationships appear when living structures must distribute forces or transport materials efficiently through tissue networks (Ball, 1999).
Structure Where Sand Breaks
Hard structure embedded in soft sediment creates pockets where currents slow and animals find shelter, turning smooth bottoms into complex habitat. | Photo credit: A. Mitchell
Beneath the surface where oyster shells, coquina fragments, and storm-scattered debris interrupt the sand, the bottom shifts from smooth sediment to broken relief. In these pockets of structure, octopuses occupy cavities narrow enough to seal with the mantle.
Field observations show that octopus dens occur most frequently within crevice-rich substrates where structural complexity provides refuge and leverage for movement and defense (Anderson et al., 2002). Small fish hover near the edges of these openings, maintaining circular perimeters that expand and contract with the reach of a hidden arm. Juvenile sheepshead pick along shell ridges in repeating passes, their feeding paths tracing arcs that mirror the curvature of the structure beneath them.
Within these shelters, the eight arms of an octopus function as semi-independent mechanical units whose forces combine into coordinated motion (Mather & O’Dor, 1991). Much of this control occurs locally within the arms themselves, allowing rapid adjustment as the animal navigates complex surfaces.
As currents pass through these cavities, suspended particles settle into protected depressions, feeding microbial films that alter oxygen exchange and nutrient cycling along the bottom boundary. Structural geometry therefore governs not only animal behavior but also the micro-distribution of material across the seafloor.
Spirals Carried Offshore
As a thin sheet of water drains across the sand, it splits into branching paths that curve and merge before disappearing. These temporary channels briefly record how moving water redistributes energy along the shoreline. | Photo credit: A. Mitchell
Outside the inlet bars, a drifting boat leaves a wake that separates into tightening vortices. Each eddy contracts as it rotates, conserving angular momentum while turbulence redistributes energy through surrounding water.
Similar rotating structures form within rip currents, where narrow jets of water moving seaward generate circulation cells that trap plankton and suspended particles (Feddersen, 2014; MacMahan et al., 2006; Thorpe, 2005).
Fluid motion often organizes into spiraling paths under these conditions, reflecting the conservation of momentum within rotating systems (Longuet-Higgins, 1969; Peregrine, 1976).
Foam left behind by receding breakers sometimes curls into arcs that briefly echo shell fragments scattered across the wash.
Schools of baitfish caught at the margins of these rotations may briefly organize into crescent formations before the structure dissolves.
Incoming waves arrive in layered packets because slightly offset frequencies overlap and reinforce one another. When multiple rhythms travel through the same body of water, their interaction produces envelopes of larger motion surrounding smaller oscillations (Longuet-Higgins, 1969).
From the deck of a small boat these envelopes pass as broad rises containing finer pulses, a hierarchy of motion that continuously reshapes sandbars and sediment pathways along the coast.
Circles the Water Keeps
A fish briefly touching the surface sends expanding rings across the water, one of the simplest expressions of circular motion in nature. | Photo credit: A. Mitchell
At creek mouths and along nearshore bars, circles appear and vanish faster than the eye can catalogue them. These expanding rings are among the simplest patterns in nature, appearing whenever energy spreads outward through still water.
A ripple expands from a falling drop, its edge widening until it meets another wave and dissolves into interference. The distance around that circle always exceeds the span across it by the same proportion — the constant mathematicians call π.
Circular motion governs more than surface ripples. Tidal creeks bend into loops where erosion and sediment deposition redistribute its momentum along the channel edges that gradually produce curved meanders (Phillips, 1977; Temmerman et al., 2007; Seminara, 2006).
Within these bends, suspended sediment slows and settles, forming point bars that redirect flow during the next tidal cycle.
Offshore, rotating eddies may close into temporary rings that trap plankton and organic particles before dissolving again (MacMahan et al., 2006).
The circle becomes a moving boundary that regulates exchange while it lasts.
Proportion in a Moving Margin
Sunlight reflecting across shallow ripples reveals the repeating wave patterns that constantly reshape coastal sand flats. | Photo credit: A. Mitchell
Across marsh edge, wrack line, and nearshore water, similar patterns recur because natural systems governed by energy exchange tend to converge toward stable configurations.
Spiral drainage, fivefold symmetry, clustered leaf spacing, rotating vortices, and circular ripples represent different expressions of the same negotiation between force and structure.
Across biological and physical systems, recurring proportional relationships often emerge because they minimize energetic cost while maintaining stability (Ball, 1999; Cross & Hohenberg, 1993; Rodriguez-Iturbe & Rinaldo, 1998).
As sediment accumulates or erodes and vegetation thickens or thins, these geometric tendencies alter water residence time, root exposure, and nutrient retention within the marsh (Fagherazzi et al., 2013).
Each tide crosses the boundary again.
And each time it does, the coast recalculates its proportions.
References
Anderson, R. C., Wood, J. B., & Byrne, R. A. (2002). Octopus senescence: The beginning of the end. Journal of Applied Animal Welfare Science, 5(4), 275-283. https://doi.org/10.1207/s15327604jaws0504_02
Beadle, S. C. (1989). Ontogenetic regulatory mechanisms, heterochrony, and eccentricity in dendrasterid sand dollars. Paleobiology, 15(3), 205-222. https://doi.org/10.1017/s0094837300009428
Bouma, T. J., Friedrichs, M., Van Wesenbeeck, B. K., Temmerman, S., Graf, G., & Herman, P. M. (2009). Density‐dependent linkage of scale‐dependent feedbacks: A flume study on the intertidal macrophyte Spartina anglica. Oikos, 118(2), 260-268. https://doi.org/10.1111/j.1600-0706.2008.16892.x
Chan, B. K., Chan, K. K., & Leung, P. C. (2006). Burrow architecture of the ghost crab Ocypode ceratophthalma on a sandy shore in Hong Kong. Hydrobiologia, 560(1), 43-49. https://doi.org/10.1007/s10750-005-1088-2
Cross, M. C., & Hohenberg, P. C. (1993). Pattern formation outside of equilibrium. Reviews of Modern Physics, 65(3), 851-1112. https://doi.org/10.1103/revmodphys.65.851
Douady, S., & Couder, Y. (1996). Phyllotaxis as a dynamical self organizing process part II: The spontaneous formation of a periodicity and the coexistence of spiral and whorled patterns. Journal of Theoretical Biology, 178(3), 275-294. https://doi.org/10.1006/jtbi.1996.0025
Fagherazzi, S., Mariotti, G., Wiberg, P., & McGlathery, K. (2013). Marsh collapse does not require sea level rise. Oceanography, 26(3), 70-77. https://doi.org/10.5670/oceanog.2013.47
Feddersen, F. (2014). The generation of Surfzone eddies in a strong alongshore current. Journal of Physical Oceanography, 44(2), 600-617. https://doi.org/10.1175/jpo-d-13-051.1
Kirwan, M. L., & Murray, A. B. (2007). A coupled geomorphic and ecological model of tidal marsh evolution. Proceedings of the National Academy of Sciences, 104(15), 6118-6122. https://doi.org/10.1073/pnas.0700958104
Leonard, L. A., & Luther, M. E. (1995). Flow hydrodynamics in tidal marsh canopies. Limnology and Oceanography, 40(8), 1474-1484. https://doi.org/10.4319/lo.1995.40.8.1474
Lim, S. S., Tantichodok, P., & Yong, A. Y. (2011). Comparison of burrow morphology of juvenile and young adult Ocypode ceratophthalmus from sai Kaew, Thailand. Journal of Crustacean Biology, 31(1), 59-65. https://doi.org/10.1651/10-3314.1
Longuet-Higgens, M. S. (1969). On the joint distribution of wave periods and heights. Journal of Marine Research, 27, 1-16.
Lucrezi, S., Schlacher, T. A., & Walker, S. (2009). Monitoring human impacts on sandy shore ecosystems: A test of ghost crabs (Ocypode spp.) as biological indicators on an urban beach. Environmental Monitoring and Assessment, 152(1-4), 413-424. https://doi.org/10.1007/s10661-008-0326-2
Mather, J., & O’Dor, R. (1991). Foraging strategies and predation risk shape the natural history of juvenile Octopus vulgaris. Bulletin of Marine Science, 49(1-2), 256-269.
Mooi, R., & David, B. (1998). Evolution within a bizarre phylum: Homologies of the first echinoderms. American Zoologist, 38(6), 965-974. https://doi.org/10.1093/icb/38.6.965
Murray, A. B., & Paola, C. (1994). A cellular model of braided rivers. Nature, 371(6492), 54-57. https://doi.org/10.1038/371054a0
Philips, O. M. (1977). The dynamics of the upper ocean (2nd ed.). Cambridge University Press.
Rodriguez-Iturbe, I., Rinaldo, A., & Levy, O. (1998). Fractal river basins: Chance and self-organization. Physics Today, 51(7), 70-71. https://doi.org/10.1063/1.882305
Seminara, G. (2006). Meanders. Journal of Fluid Mechanics, 554, 271-297.
Telford, M. (1981). Structural analysis of the test of echinoids. Zoomorphology, 98, 93-110.
Temmerman, S., Bouma, T., Van de Koppel, J., Van der Wal, D., De Vries, M., & Herman, P. (2007). Vegetation causes channel erosion in a tidal landscape. Geology, 35(7), 631. https://doi.org/10.1130/g23502a.1
Several species of jellyfish appear along the waters of Onslow County, North Carolina as the coastal ecosystem shifts from winter toward spring. Moon jellies, comb jellies, sea nettles, and cannonball jellyfish all move through these waters at different times of year, responding to temperature, tides, and the seasonal return of plankton (Purcell et al., 2007; Lucas et al., 2012; Cloern & Jassby, 2010).
At the Edge of Winter
Late winter along the estuarine marshes of Onslow County. Marsh grasses remain the color of dried straw while the coastal ecosystem waits for spring. | Photo credit: M. Mitchell
In late February along the Intracoastal Waterway, the coast exists in a kind of suspension. The marsh grasses behind Topsail Island are still the color of dried straw, their green not yet returned. The wind carries more memory than warmth, and the water — though brighter in the lengthening light — remains clear in the way cold water often is, revealing sandy bottom, oyster shell, and shadow without the haze of summer plankton.
Nothing looks abundant. Nothing appears urgent. The shoreline feels patient.
If you lean over a dock and allow your eyes to adjust, the surface begins to resolve into layers. What first appears empty reveals movement — a faint pulse beneath the water, nearly invisible unless sunlight strikes at the right angle. A small translucent bell, no wider than your palm, opens and closes in a steady rhythm while the current carries it sideways through the creek.
The first jellies of the season are easy to miss.
They are small. They are clear. And they belong to the quiet phase of the coastal year.
The Water Before Summer
Early spring estuarine water along the coast often appears clear and quiet as plankton populations begin rebuilding after winter. | Photo credit: A. Mitchell
Early spring water along the southern North Carolina coast often carries a glass-like quality. Plankton populations are rebuilding after winter. Suspended sediments have settled during calmer stretches. Against that clarity, the earliest gelatinous drifters seem almost designed to disappear.
Standing along the docks and creeks of Onslow County, most of what we notice are the drifting bells moving slowly through the water.
But the life of a jellyfish does not begin there.
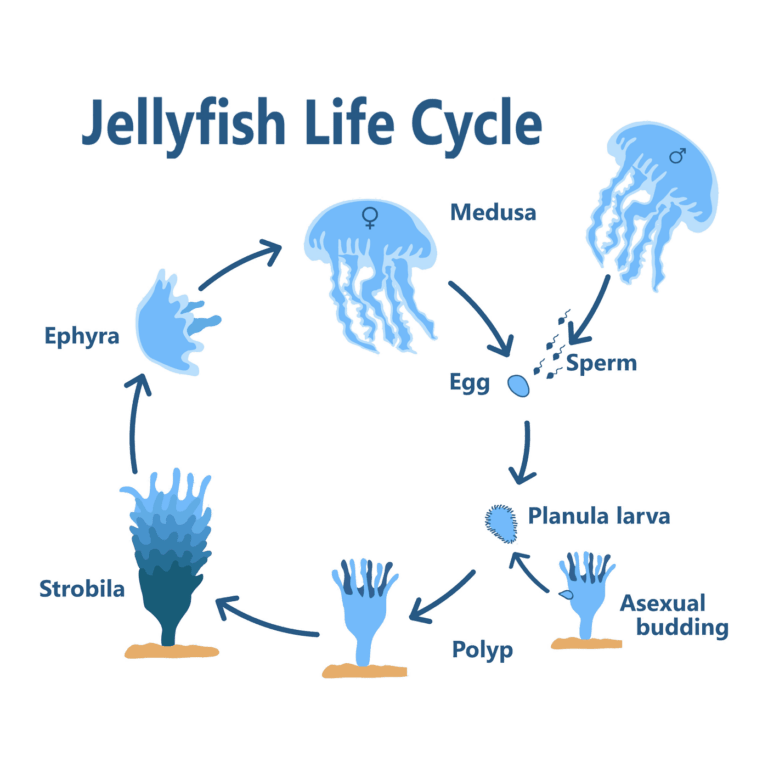
How a Jellyfish Begins
The jellyfish most people recognize — the drifting bell, the trailing tentacles — is only one phase of a much longer life cycle.
Generalized life cycle of a true jellyfish. Many species begin as microscopic larvae that settle into tiny polyps attached to submerged surfaces before releasing young jellyfish into the water column. | Graphic credit: Key West Aquarium
Most true jellyfish begin as fertilized eggs released into the water column. Each egg develops into a tiny, free-swimming larva called a planula. There are many planulae in the water at once, but each one is a single organism, smaller than a grain of sand, carried by currents you would never notice from the surface.
Within a few days — sometimes less than a week, depending on temperature — a planula settles onto a hard surface. It may attach to the underside of a dock piling, the rough edge of an oyster shell, a shaded bridge support, or even a shell resting quietly in the mud. Once attached, it transforms into a polyp (Lucas et al., 2012).
In this stage it does not resemble a jellyfish at all. It is small — often only a few millimeters tall — no larger than a grain of rice. If you could flip that piling into the sunlight in February, you would not see a jellyfish.
You would see something that looked more like a pale freckle against the wood.
And yet, that freckle holds potential.
The polyp may remain in that form for months. Anchored beneath docks and along oyster beds, it feeds on microscopic prey drifting past and survives the colder stretch of winter water (Purcell, 2012).
That freckle — that rice-sized polyp — does not always remain alone.
In some species of true jellyfish, the polyp stage can reproduce asexually, forming small copies of itself along the same piling or oyster shell (Purcell, 2012).
When spring begins to soften the creek and temperatures rise, the polyp changes again. In a process known as strobilation, its body reorganizes into stacked segments (Purcell et al., 2007; Lucas et al., 2012). One by one, those segments separate into the water as tiny juvenile jellyfish called ephyrae (Purcell et al., 2007).
Newly released ephyrae — juvenile jellyfish — pulsing through the water after separating from the polyp stage during strobilation.
As the ephyra develops, its arms fill in and smooth into a rounded bell. It becomes the drifting medusa we recognize along docks, tidal creeks, and open shoreline.
As the ephyra develops, its arms fill in and smooth into a rounded bell. It becomes the drifting medusa we recognize along docks, tidal creeks, and open shoreline.
If you step away from the dock and look toward the darker center of the creek, the pattern shifts without breaking. Not every gelatinous drifter begins life attached to wood or shell. Comb jellies live their entire lives suspended in open water, developing and reproducing where freshwater flowing downriver meets saltwater moving inland on the tide (Purcell et al., 2001).
What can seem like separate coastal experiences — the pale freckle beneath a dock, the first small jellies of spring, the sudden sting beneath a swimsuit — are phases of the same unfolding life cycle.
A Life Without a Brain
Watch a jellyfish long enough drifting beneath a dock or through the calm water of a tidal creek in Onslow County, and the question eventually arises.
What is directing it?
The bell contracts. The animal pulses forward. Tentacles drift outward and close around passing prey. The movement appears deliberate, almost rhythmic, as though some quiet decision were being made.
And yet, jellyfish have no brain.
Instead, their bodies are organized around a diffuse network of nerve cells known as a nerve net. Sensory information travels across this web of neurons distributed throughout the bell and tentacles — more like signals moving along a strand of Christmas lights, where each bulb responds along the line, rather than a single switch controlling everything at once (Mackie & Meech, 1995).
Moon jellyfish pulsing through the water. Jellyfish lack a centralized brain; instead, a diffuse nerve net distributed throughout the bell coordinates their movement and responses to the surrounding water.
Along the margin of the bell, specialized sensory structures known as rhopalia help them sense orientation and balance in the water column. In a way, they function a bit like the balance sensor in a phone that knows when the screen should rotate (Garm et al., 2006; Skogh et al., 2006).
None of these signals pass through a central command center.
Instead, the entire body participates in sensing the surrounding water.
The current shifts. Light filters through the surface. Something brushes against the tentacles.
And the jelly responds.
Moon Jellies: The Quiet Pulsers Near Structure
A moon jelly drifting beneath a dock in early spring may be only a few inches across. Its bell is nearly colorless, soft at the edges, its body so transparent that it seems less like an animal and more like a moving lens in the water.
What often gives it away are four faint circles inside the bell — pale rings that resemble small moons suspended within the jelly. Those structures are reproductive organs, and they are the feature that gives the species its common name.
Moon jellyfish (Aurelia aurita) along the shoreline. The four horseshoe-shaped structures visible inside the bell are reproductive organs — the feature that gives the species its “moon jelly” name. | Photo credit: oosty, iNaturalist
Around the margin of the bell hang delicate, hair-fine tentacles. They are far shorter and less conspicuous than the trailing threads of sea nettles that appear later in the summer.
Earlier in its life it exists as a tiny polyp attached beneath docks and oyster shells — the same pale “freckles” that persist quietly through the winter on the shaded structures below the waterline.
As the water slowly warms into the upper 40s and 50s °F (8–13°C), those anchored moon jelly polyps begin releasing young jellyfish into the creek in a process scientists call strobilation (Purcell et al., 2007; Purcell, 2012).
As these young jellies drift through the creek, their bells pulse slowly against the current. The water around them carries clouds of microscopic life — copepods and other plankton rebuilding after winter — and whatever brushes the tentacles becomes food (Lucas et al., 2012; Cloern & Jassby, 2010).
Their tentacles do carry stinging cells, called nematocysts, like microscopic harpoons built to capture animals far smaller than we are. For most people, those harpoons are too small to penetrate the outer layer of human skin.
A swimmer may brush past a moon jelly without feeling anything at all.
Comb Jellies: The Invisible Drifters of Open Water
Comb jellies — ctenophores such as Mnemiopsis leidyi — are even more elusive.
They lack stinging cells and instead capture prey with sticky cells (Purcell et al., 2001). Their bodies are almost entirely transparent. What gives them away are rows of tiny beating cilia that catch the light and flash briefly like moving prisms.
In darkness, some comb jellies can also produce brief flashes of bioluminescent light when disturbed, though along the creeks of Onslow County what we usually notice are the shifting rainbows created as sunlight bends through their beating cilia.
Comb jelly (ctenophore) drifting in the water column. Unlike true jellyfish, comb jellies lack stinging cells and capture prey using sticky cells. | Photo credit: A. Mitchell
Unlike moon jellies that may first appear near docks and pilings, comb jellies are often more noticeable in slightly deeper portions of tidal creeks and open estuary as spring advances (Purcell et al., 2001).
From the dock, they can be invisible.
But scoop a bucket of water in late spring or early summer and the illusion changes. What looked like empty creek water suddenly fills with small gelatinous spheres — clear, bead-like forms tumbling gently against one another, not unlike the soft water beads children play with, often called Orbeez.
In a net, they resemble scattered jelly stones.
They have been there all along.
The difference is scale and perspective.
Sea Lice: The Unseen Larvae of Warm Days
By late spring another gelatinous presence begins to make itself known, though most people never see the organism responsible.
On calm, warm days along the beach, swimmers sometimes step from the water with a faint prickling sensation along their skin. The irritation may begin around the ankles, between the toes, or beneath a swimsuit where fabric presses against the body. Hours later a rash can appear.
Locally this irritation is often called “sea lice,” though the name is misleading. They are not lice at all. The sensation comes from microscopic cnidarians — most commonly the larval stages of the thimble jellyfish (Linuche unguiculata), though larvae of certain sea anemones can produce the same reaction (Segura-Puertas et al., 2001; Wong et al., 1994).
Thimble jellyfish (Linuche unguiculata). The microscopic larval stages of this tiny jellyfish are the most common cause of the irritation known as “sea lice,” or seabather’s eruption. | Photo credit: Foued Kaddachi
At this stage the animals are nearly invisible, drifting in the surface water. Waves and gentle onshore currents can concentrate them along the shoreline, the same shallow areas where swimmers enter the water, children play in the surf, and beachgoers wade while searching for shells.
When these larvae become trapped against the skin — beneath fabric or pressed between toes and folds of skin — the same microscopic harpoons, or nematocysts, used to capture prey can inject a tiny amount of venom when triggered (Wong et al., 1994).
Most people recognize the sudden prickling sensation immediately. In the hours that follow, the irritation can intensify into a fiery rash — a reaction known medically as seabather’s eruption. Relief usually begins by rinsing the skin with fresh water after leaving the ocean and applying cold compresses to calm the irritation (Wong et al., 1994).
Few people ever see the organism responsible.
As the Season Deepens
Spring along the coast rarely arrives all at once. It unfolds in stages — water warming by degrees, plankton building slowly in the creeks and sounds, and the community of gelatinous drifters shifting with those changes.
The nearly invisible jellies of early spring give way to species that are easier to see, easier to avoid, and sometimes easier to feel.
Sea Nettles: The Summer Drifters of Brackish Water
Atlantic sea nettle (Chrysaora quinquecirrha) drifting just beneath the surface of a coastal creek. | Photo credit: A. Mitchell
By late spring and early summer, the Atlantic sea nettle (Chrysaora quinquecirrha) begins appearing more frequently in the creeks and sounds of Onslow County.
Their bells carry warm amber tones, and long tentacles trail behind them like threads drifting through the tide.
Sea nettles favor the brackish mixing zones of the estuary where freshwater flowing down the New River blends with saltwater entering through the inlets. As plankton populations increase with warming water, sea nettles follow the food supply into tidal creeks and quieter sounds (Lucas et al., 2012).
Unlike moon jellies, their nematocysts can penetrate human skin, producing the sharp sting swimmers learn to recognize.
Cannonball Jellies: The Offshore Drifters
Cannonball jellyfish (Stomolophus meleagris) found along the shoreline in Surf City, NC. Many individuals washing ashore are no longer alive once they lose the buoyant support of seawater. | Photo credit: A. Mitchell
Farther offshore another species sometimes appears — the cannonball jellyfish (Stomolophus meleagris).
Their rounded bells give them the appearance of pale drifting mushrooms or underwater buoys.
They often gather in offshore waters where ocean currents concentrate plankton (Graham et al., 2003). Storms and onshore winds can push them toward the beaches of Onslow County, where they sometimes appear along the wrack line.
Many stranded individuals are no longer alive. Their gelatinous bodies collapse quickly once they leave the buoyant support of seawater.
Beyond the breakers, however, they may still be drifting quietly through deeper currents.
An Older Pattern Beneath the Surface
It is easy to think of jellyfish as modern phenomena — summer nuisances or passing curiosities.
Yet their lineage stretches back more than 500 million years, predating vertebrates and surviving multiple mass extinctions (Cartwright et al., 2007).
Fossil impressions of ancient jellyfish alongside modern jellyfish. Soft-bodied animals like jellyfish rarely fossilize, but when preserved they reveal that jellyfish-like organisms have existed in Earth’s oceans for hundreds of millions of years. | Image credit: Fossil photo by B. Lieberman. Cunina photo by K. Raskoff, copyright.
Soft-bodied drifters pulsed through plankton-rich seas above what would eventually become Onslow County.
The small jelly beneath a dock in March is not something new.
It is continuity.
Returning to the Shoreline
Stand again at the edge of the sound as winter begins to loosen its hold on the coast.
The air is softer now. Ospreys circle overhead. Marsh grass prepares to green.
Beneath the surface, the water is changing too — warming slowly, plankton returning, currents carrying new life through the estuary.
A small bell pulses quietly past the pilings. Nearby, comb jellies flash faint rainbows when the light strikes them just right. Somewhere beyond sight, larvae drift through the tide.
None of it announces itself.
But if you lean over the water long enough in early spring, you can watch the system beginning again.
Early spring in the S. Topsail Island sound. Beneath the surface, plankton, drifting larvae, and young marine life begin to return with the warming water. | Photo credit: M. Mitchell
References
Cartwright, P., Halgedahl, S. L., Hendricks, J. R., Jarrard, R. D., Marques, A. C., Collins, A. G., & Lieberman, B. S. (2007). Exceptionally preserved jellyfishes from the Middle Cambrian. PLoS ONE, 2(10), e1121. https://doi.org/10.1371/journal.pone.0001121
Cloern, J. E., & Jassby, A. D. (2009). Patterns and scales of phytoplankton variability in estuarine–coastal ecosystems. Estuaries and Coasts, 33(2), 230-241. https://doi.org/10.1007/s12237-009-9195-3
Garm, A., Ekström, P., Boudes, M., & Nilsson, D. (2006). Rhopalia are integrated parts of the central nervous system in box jellyfish. Cell and Tissue Research, 325(2), 333-343. https://doi.org/10.1007/s00441-005-0134-8
Graham, W. M. (2001). Size-based prey selectivity and dietary shifts in the jellyfish, Aurelia aurita. Journal of Plankton Research, 23(1), 67-74. https://doi.org/10.1093/plankt/23.1.67
Graham, W. M., Pagès, F., & Hamner, W. M. (2001). A physical context for gelatinous zooplankton aggregations: A review. Jellyfish Blooms: Ecological and Societal Importance, 199-212. https://doi.org/10.1007/978-94-010-0722-1_16
Lucas, C. H., Graham, W. M., & Widmer, C. (2012). Jellyfish life histories: Role of polyps in forming and maintaining Scyphomedusa populations. Advances in Marine Biology, 133-196. https://doi.org/10.1016/b978-0-12-394282-1.00003-x
Mackie, G. O., & Meech, R. W. (1995). Central circuitry in the jellyfish Aglantha Digitale: I. The relay system. Journal of Experimental Biology, 198(11), 2261-2270. https://doi.org/10.1242/jeb.198.11.2261
Mackie, G. O., & Meech, R. W. (1995). Central circuitry in the jellyfish Aglantha Digitale: II. The ring giant and carrier systems. Journal of Experimental Biology, 198(11), 2271-2278. https://doi.org/10.1242/jeb.198.11.2271
Purcell, J. E. (2012). Jellyfish and ctenophore blooms coincide with human proliferations and environmental perturbations. Annual Review of Marine Science, 4(1), 209-235. https://doi.org/10.1146/annurev-marine-120709-142751
Purcell, J. E., Shiganova, T. A., Decker, M. B., & Houde, E. D. (2001). The ctenophore Mnemiopsis in native and exotic habitats: U.S. estuaries versus the Black Sea basin. Hydrobiologia, 451, 145-176. https://link.springer.com/article/10.1023/A:1011826618539
Purcell, J., Uye, S., & Lo, W. (2007). Anthropogenic causes of jellyfish blooms and their direct consequences for humans: A review. Marine Ecology Progress Series, 350, 153-174. https://doi.org/10.3354/meps07093
Segura-Puertas, L., Ramos, M. E., Aramburo, C., Heimer de la Cotera, E. P., & Burnett, J. W. (2001). One Linuche mystery solved: All 3 stages of the coronate scyphomedusa Linuche unguiculata cause seabather’s eruption. Journal of the American Academy of Dermatology, 44(4), 624-628. https://doi.org/10.1067/mjd.2001.112345
Skogh, C., Garm, A., Nilsson, D., & Ekström, P. (2006). Bilaterally symmetrical rhopalial nervous system of the box jellyfish Tripedalia cystophora. Journal of Morphology, 267(12), 1391-1405. https://doi.org/10.1002/jmor.10472
Wong, D. E., Meinking, T. L., Rosen, L. B., Taplin, D., Hogan, D. J., & Burnett, J. W. (1994). Seabather’s eruption: Clinical, histologic, and immunologic features. Journal of American Academy of Dermatology, 30(3), 399-406. https://www.jaad.org/article/S0190-9622(94)70046-X/abstract
Onslow County 66 Million Years Ago: Before the Coastline Existed
Today, the shoreline of Onslow County forms part of the Atlantic Coastal Plain — a broad, low landscape stretching from New Jersey to Florida (Riggs et al., 2020). Marshes, barrier islands, and estuaries define the modern coast, shaped by tides, storms, and the slow migration of sand (Riggs et al., 1995; Riggs et al., 2020). But the ground beneath those systems records a far older history. Long before marsh grass rooted the shoreline or barrier islands assembled offshore, this region lay beneath a shallow sea.
Reconstruction of North America during the Late Cretaceous, when high global sea levels flooded large portions of the continent. Shallow epicontinental seas covered much of the interior and extended across the Atlantic Coastal Plain, submerging regions that would later become the southeastern United States. | Image credit: Illustration of Late Cretaceous North America showing the Western Interior Seaway. Adapted from U.S. Geological Survey educational materials.
Onslow County 66 million years ago looked nothing like it does today. If you could stand where the county sits at the end of the Cretaceous, there would be no ground beneath your feet. No marsh, no barrier islands, no inlet channels breathing with tide. The coastline lay far inland. Eastern North Carolina was submerged beneath a warm, shallow sea that stretched in a broad, quiet shelf from the continent toward an ocean still reorganizing after the breakup of Pangaea.
If a human body could enter that sea, the first sensation would not be clarity but thickness. The water would feel warm and heavy against skin, dense with suspended life. Visibility would shorten to a green haze where light diffused instead of traveling cleanly. Through that haze, movement would register before shape: the passage of large animals built for a shelf no longer occupied by their kind — mosasaurs turning in slow arcs, plesiosaurs rising through layered water, sharks, already ancient, tracing patrol routes beneath them. Each body would displace the plankton-rich column, sending pressure outward. The ocean would feel less like empty space and more like a corridor constantly shared. Each movement would push through plankton-rich water that behaved less like modern surf and more like a living suspension, as if the ocean itself carried weight.
The water would have been green with plankton, heavy with suspended carbonate, the kind of sea that builds geology slowly from drifting skeletons. There were no beaches yet because there was no edge — only a gradual transition from submerged coastal plain into open Atlantic. The sediments beneath modern Onslow County record this as stacked marine layers: sand, clay, marl, and chalky limestones built from microscopic shells settling through millions of seasons (Miller et al., 2005).
This was not an empty sea. It was structured like a city.
Reef communities rose from carbonate platforms. Ammonites spiraled through open water. Early teleost fishes filled midwater niches (Friedman, 2010; Near et al., 2013). Marine reptiles — mosasaurs and plesiosaurs — patrolled the upper food web. And sharks, already ancient by this point, occupied ecological roles recognizable even now: cruisers of the shelf, opportunists of the drop-off, specialists shaped by tooth and speed.
The modern Atlantic Coastal Plain is the memory of that sea compressed into stone.
Teeth as Geological Fossils
Fossil tooth of the giant shark, Otodus megalodon, found in the surf at Surf City, North Carolina. Teeth like this erode from Miocene and Pliocene marine sediments beneath the Atlantic Coastal Plain, remnants of an ocean that covered this region millions of years ago. | Photo credit: Alicia Sanders, 2025
The shark teeth found along Onslow beaches are not simply remnants of animals; they are fragments of sedimentary history washing back to the surface. Rivers, storms, dredging, and shoreline erosion re-expose marine layers that were buried when sea level fell and the continent emerged. Each tooth has traveled twice: first through the animal that grew it, and later through millions of years of burial, erosion, and exposure.
Many of the large triangular teeth people call “megalodon” are younger than the Cretaceous itself. The giant shark Otodus megalodon lived much later, during the Miocene and Pliocene, roughly 23 to 3.6 million years ago — a reminder that the Atlantic shelf has been a marine environment repeatedly across deep time (Pimiento & Balk, 2015). The reason both Cretaceous and Miocene fossils appear in the same coastal region is not contradiction but layering. Eastern North Carolina is a staircase of ancient seas, each episode leaving deposits that modern erosion cross-cuts and reveals.
The shoreline acts like a rotating archive. Storms turn the pages.
The Cretaceous: A Climate Without Ice
Marine reptiles dominated ocean food webs during the Mesozoic Era, when dinosaurs ruled on land. Mosasaurs and other large predators hunted fish, ammonites, and other marine animals in the warm seas that covered much of North America. | Illustration credit: Smithsonian Institution
The Cretaceous sea covering Onslow County existed in a greenhouse world. There were no polar ice caps. Global temperatures were higher. Sea levels stood among the highest in the last 500 million years. Warm currents circulated freely between basins, and the shallow epicontinental seas were engines of biodiversity (Hay, 2011).
In such climates, coastal ecosystems functioned differently. Productivity was driven by ocean circulation and nutrient upwelling rather than the strong seasonal temperature swings that structure many modern coastal systems. Carbonate production accelerated. Marine food webs expanded vertically, filling ecological space with specialists. The shallow shelf that covered North Carolina would have been biologically dense — a continuous gradient from estuarine margins to open marine habitats, without the sharp land–sea boundary we recognize today.
The modern Outer Banks, in this sense, are a recent invention. They are sand arranged by late-Quaternary sea-level oscillation. The deeper story of this coast is marine.
Extinction as a Geological Boundary
At the end of the Cretaceous, about 66 million years ago, the asteroid impact now known as the Chicxulub event closed this chapter abruptly. Marine ecosystems did not vanish overnight; they reorganized under the cascading collapse of planktonic food webs (Schulte et al., 2010). The sedimentary record along the Atlantic margin preserves this boundary as a thin horizon enriched in iridium — a planetary fingerprint marking a moment when global systems reset.
A rock sample showing the Cretaceous–Paleogene boundary, where a thin iridium-rich layer marks the global extinction event triggered by the Chicxulub asteroid impact about 66 million years ago. Specimen from Wyoming, displayed at the San Diego Natural History Museum.| Photo credit: Fossil Crates, 2022
In the hundreds of thousands to millions of years that followed, ocean ecosystems slowly rebuilt (Schulte et al., 2010; Friedman, 2010). Plankton communities recovered first, allowing marine food webs to reassemble from the bottom upward.
The sea that covered Onslow County withdrew gradually over the tens of millions of years following the Cretaceous–Paleogene boundary, not because of the impact alone, but because tectonics and climate redirected Earth’s balance of water and land. Through the Paleogene and into the Neogene, regression exposed portions of the Atlantic Coastal Plain. Rivers carved channels into former seabeds. Marshes colonized low ground. Much later, during the Pleistocene ice-age cycles beginning about 2.6 million years ago, barrier islands assembled from mobile sand as sea level rose and fell repeatedly.
What we walk today is the lifted floor of a vanished ocean.
This interactive reconstruction shows how Earth’s continents shifted through deep time. At 66 million years ago, much of the Atlantic Coastal Plain—including modern eastern North Carolina—lay beneath a warm shallow sea. | Visualization credit: Ian Webster, dinosaurpictures.org
The end of the Cretaceous did not simply erase species; it reorganized the architecture of marine life. In the first several hundred thousand years after the Chicxulub impact, the collapse of plankton communities removed the base of food webs that had supported ammonites, many marine reptiles, and numerous large predatory fishes. Apex niches did not stay empty for long. During the early Paleogene, roughly 66 to 50 million years ago, sharks, teleost fishes, and early marine mammals diversified rapidly into the ecological space left behind (Schulte et al., 2010; Friedman, 2012).
Reconstruction of marine life near the end of the Cretaceous Period, shortly before the Chicxulub asteroid impact triggered a global mass extinction about 66 million years ago. The event collapsed marine food webs and reshaped ocean ecosystems across the planet. | Illustration: Smithsonian Institution
In the aftermath, the shelf would not look empty at first glance but would feel altered, as large bodies that once displaced water in constant motion were absent, leaving the vertical space above the seafloor open, quieter, and less crowded. A swimmer would sense the difference not through sight alone but through the water’s stillness — fewer passing pressure waves and fewer shadows interrupting the light.
In the shallow seas that once covered eastern North Carolina, this transition marked a shift from reptile-dominated predator guilds to fish- and shark-centered systems. Survivors tended to share traits that remain advantageous in modern estuaries: flexible diets, rapid reproduction, and tolerance for fluctuating conditions. The extinction boundary favored generalists over specialists, and lineages capable of exploiting disrupted ecosystems seeded the foundation of the modern Atlantic marine fauna.
The Cretaceous sea did not end — it evolved under constraint.
The Miocene: A Predator-Rich Shelf
Over the tens of millions of years that followed the Paleogene recovery, marine ecosystems continued diversifying as continents drifted toward their modern positions and ocean circulation strengthened. By the Miocene, roughly 23 to 5 million years ago, the sea covering eastern North Carolina was not the same water body that drowned the Cretaceous coast. Continents had shifted. Currents reorganized. The Atlantic margin was beginning to resemble its modern geometry. What remained constant was the shelf: shallow, warm, nutrient-rich, and biologically crowded.
The Miocene shelf was structured by productivity. Warm global climates intensified circulation patterns that mixed nutrients and supported dense prey fields. Where energy concentrates, ecosystems scale upward. Plankton blooms fueled vast schools of fish and squid that moved through the water column in shifting layers (Hay, 2012; Pimiento et al., 2016).
Reconstruction of a Miocene marine ecosystem (~6 million years ago) featuring dolphins (Eurhinodelphis), a penguin (Spheniscus), the long-necked seal Acrophoca, and the giant shark, Otodus megalodon. During the Miocene, the diversification of marine mammals supported some of the largest predators in ocean history. | Artwork by Julius Csotonyi
This abundance supported a growing diversity of marine vertebrates (Pimiento et al., 2016). Whales diversified explosively: early baleen whales filtered plankton blooms that pulsed across the shelf, while toothed whales pursued schooling fish and squid, their passage shifting the light before their bodies came fully into view. Pinnipeds hauled out on emergent islands. Sea turtles nested along coastlines that advanced and retreated with slow tectonic breathing. Below that movement, sirenian grazers — ancestors of modern manatees — moved slowly through seagrass beds, shaping the shelf from the bottom while predators ruled above (Domning, 2001).
To occupy that Miocene shelf as a small observer would be to feel scale in motion. Migrating whales and large predators would load the surrounding water with momentum before they arrived, a pressure you could feel before you could see its source. The shelf seemed to flex around movement, the water itself shaped by the animals traveling through it. The water carried that weight differently than in the Cretaceous — not the suspension of reptile-dominated seas, but the mass of mammals built for speed and scale.
In ecosystems where prey concentrates and marine mammals flourish, apex predators inevitably emerge. In the Miocene Atlantic, that role belonged to the giant shark whose teeth still surface along North Carolina beaches: Otodus megalodon. Its immense size was not evolutionary extravagance but ecological arithmetic. A predator of that scale can exist only where the energy flowing through the system is great enough to sustain it (Pimiento et al., 2016).
Sediments from Miocene deposits in the Atlantic Coastal Plain preserve a fossil record dominated by marine vertebrates. These fossils accumulate in the same geological staircase as older layers, which is why storms today liberate teeth from multiple epochs simultaneously. The shoreline is a cross-section through predator history.
Megalodon disappears near the Pliocene boundary, likely a casualty of ecological restructuring — shrinking nursery habitat, prey redistribution, and competition from emerging marine mammals (Pimiento & Clements, 2014). The predator city did not vanish; it reorganized.
The Pleistocene: Ice, Sand, and a Moving Coast (~2.6 million – 11,700 years ago)
During glacial low stands, standing on the exposed shelf would produce a disorienting absence. Wind would move across ground that remembered being ocean. The surface would hold the texture of former seabed — compacted, rippled, cut by channels where rivers extended into newly revealed terrain. Air would replace water pressure, but the land would still read as marine, a coastline temporarily paused in withdrawal.
The Pleistocene introduced a rhythm that still governs the modern coastline: glacial cycling. Ice sheets expanded and retreated dozens of times, locking ocean water onto continents and then releasing it. Each cycle shifted sea level by tens of meters. Eastern North Carolina repeatedly alternated between exposed coastal plain and submerged shelf (Lambeck et al., 2014).
When sea levels fell, rivers carved deeply into former seabeds, cutting channels that later became estuaries, while rising seas flooded those valleys again, redistributing sand along migrating shorelines as barrier islands assembled from sediment sorted into long, mobile ridges by waves and currents (Riggs et al., 1995; Riggs et al., 2020).
Reconstruction of a shallow coastal ecosystem similar to those that developed along the Atlantic Coastal Plain as sea levels stabilized after the Ice Age. Seagrass beds, shellfish, fishes, turtles, and marine mammals formed the foundation of the modern estuarine communities that now define the Carolina coast. | Painting by Michael Rothman, Florida Museum of Natural History.
The coast stopped being a static shelf and became a machine in motion, and Pleistocene ecosystems were shaped by that instability as species adapted to shifting salinity, temperature, and shoreline position. Many cold-adapted megafauna disappeared or shifted poleward, while warm-temperate estuarine assemblages consolidated in their place.
The ecological winners were organisms capable of building habitat: marsh grasses trapping sediment, oysters engineering reefs, and filter feeders clarifying water. Those assemblages increasingly resembled the modern Atlantic shelf, with drum, croaker, and mullet occupying estuarine corridors carved by drowned rivers while rays and small coastal sharks patrolled nursery shallows. Oyster reefs rose in dense clusters, and early marsh communities anchored sediment with grasses similar to those that now define the Carolina coastline.
The emerging system favored species tolerant of fluctuation — animals able to move with the shoreline rather than resist it — as cooling climates and destabilized coasts increasingly selected for flexibility over scale, replacing the giants of the Miocene shelf with communities built for movement rather than permanence.
The modern Onslow estuary is therefore a recent equilibrium layered atop instability.
Pleistocene Transition: From Ice Age Coast to Modern Estuary
Shifts in the eastern North American coastline from the Last Glacial Maximum (~20,000 years ago) to the present. During the Ice Age, much of the continental shelf was exposed land. As glaciers melted and sea level rose, coastal rivers flooded and estuaries formed, creating the modern Atlantic Coastal Plain shoreline.| Illustration credit: USGS, Water Science School
The close of the Pleistocene did not feature a single catastrophic boundary but a climatic stabilization. As the last major ice sheets retreated about 11,700 years ago, sea level rose rapidly and then slowed. Coastlines stopped migrating at glacial speed. Estuaries stabilized long enough for persistent marsh systems to develop. Oyster reefs expanded. Seagrass beds colonized shallow bays (Lambeck et al., 2014; Kennett & Shackleton, 1975).
The organisms that dominate modern estuaries are ecosystem engineers. They do not simply inhabit the coast — they build it.
What appears ancient in the marsh is, in geological terms, newly assembled.
Survivors in Motion: Sharks and Teleost Continuity
Standing in modern surf, that continuity is still tactile. The water along the shelf carries suspended sand and organic haze, softening visibility to a few body lengths. Something large can pass nearby without breaking the surface, announced only by a shift in current or a vibration through the feet. The present ocean feels busy in the same quiet way ancient shelves must have felt — full of motion just beyond clear sight. The modern shelf feels lighter, but not empty. The water still carries motion long before form appears, transmitting the passage of sandbar sharks, blacktips, and schooling menhaden through vibration rather than sight. The weight is subtler now — distributed across smaller bodies, faster cycles, suspended sand and organic haze — yet the sensation remains – a medium that remembers being crowded.
The most striking feature of the Atlantic shelf is not how much has changed, but how much has endured. Sharks were already ancient when the Cretaceous sea covered this region, their lineage extending back more than 400 million years (Ebert et al., 2021).
Ancient oceans and modern seas share the same ecological architecture. Slide to compare a Late Cretaceous shark community with a modern marine ecosystem. While species have changed across millions of years, sharks and teleost fishes continue to occupy many of the same roles within ocean food webs. | Left image: Late Cretaceous marine assemblage
Modern coastal sharks reflect this inheritance, with sandbars, blacktips, bonnetheads, and dogfish representing lineages refined through repeated ecological resets (Ebert et al., 2005). Teleost fishes tell a parallel story: after the end-Cretaceous extinction, they diversified explosively, filling feeding niches that define modern marine communities (Friedman, 2010).
Species change across epochs, but functional roles persist, and the system remembers its architecture even when its cast rotates. The sharks offshore now are not echoes of a lost world; they are its direct continuation.
The Coast as a Layered Archive
Shell fragments scattered across the shoreline near Surf City, North Carolina. Each tide exposes pieces of past marine life, reminders that the modern coast sits atop layers of older oceans and ecosystems.| Photo credit: Tom’s Teeth, 2019
When a fossil tooth surfaces in the surf along the Onslow County coast, it is not emerging from a single time but from stacked histories compressed beneath the modern shoreline. Cretaceous seas. Miocene predator guilds. Pleistocene shorelines advancing and retreating with ice age pulses. Each episode writes a layer. Storm energy and human dredging occasionally cut into those layers, returning fragments to circulation.
This is why the coast feels haunted by deep time. The sediment is not just sand; it is a palimpsest of ecosystems.
The sharks that swim offshore now — sandbars, blacktips, bonnetheads — are heirs to lineages that survived the extinction boundary and adapted through cycles of climate and geography. Their teeth will enter the archive in turn. Millions of years from now, another shoreline will release them, and a different species will walk a beach made from our present seafloor.
The coast is not a place fixed in space. It is a moving edge between worlds, carrying memory forward grain by grain.
Domning, D. P. (2001). The earliest known fully quadrupedal sirenian. Nature, 413(6856), 625-627. https://doi.org/10.1038/35098072
Ebert, D. A., Dando, M., & Fowler, S. (2021). Sharks of the world: A complete guide. Princeton University Press.
Friedman, M. (2010). Explosive morphological diversification of spiny-finned teleost fishes in the aftermath of the end-Cretaceous extinction. Proceedings of the Royal Society B: Biological Sciences, 277(1688), 1675-1683. https://doi.org/10.1098/rspb.2009.2177
Hay, W. W. (2012). Experimenting on a small planet: A scholarly entertainment (1st ed.). Springer Science & Business Media.
Kennett, J. P., & Shackleton, N. J. (1975). Laurentide ice sheet Meltwater recorded in Gulf of Mexico deep-sea cores. Science, 188(4184), 147-150. https://doi.org/10.1126/science.188.4184.147
Lambeck, K., Rouby, H., Purcell, A., Sun, Y., & Sambridge, M. (2014). Sea level and global ice volumes from the last glacial maximum to the Holocene. Proceedings of the National Academy of Sciences, 111(43), 15296-15303. https://doi.org/10.1073/pnas.1411762111
Miller, K. G., Kominz, M. A., Browning, J. V., Wright, J. D., Mountain, G. S., Katz, M. E., Sugarman, P. J., Cramer, B. S., Christie-Blick, N., & Pekar, S. F. (2005). The Phanerozoic record of global sea-level change. Science, 310(5752), 1293-1298. https://doi.org/10.1126/science.1116412
Near, T. J., Dornburg, A., Eytan, R. I., Keck, B. P., Smith, W. L., Kuhn, K. L., Moore, J. A., Price, S. A., Burbrink, F. T., Friedman, M., & Wainwright, P. C. (2013). Phylogeny and tempo of diversification in the superradiation of spiny-rayed fishes. Proceedings of the National Academy of Sciences, 110(31), 12738-12743. https://doi.org/10.1073/pnas.1304661110
Pimiento, C., Balk, M., & Celements, C. (2014). Reconstructing the extinction of the giant Megalodon shark (Carcharcoles Megalodon). The Paleontological Society Special Publications, 13, 52-52. https://doi.org/10.1017/s2475262200011102
Pimiento, C., & Balk, M. A. (2015). Body-size trends of the extinct giant shark Carcharocles megalodon : A deep-time perspective on marine APEX predators. Paleobiology, 41(3), 479-490. https://doi.org/10.1017/pab.2015.16
Pimiento, C., & Clements, C. F. (2014). When did Carcharocles megalodon become extinct? A new analysis of the fossil record. PLoS ONE, 9(10), e111086. https://doi.org/10.1371/journal.pone.0111086
Pimiento, C., MacFadden, B. J., Clements, C. F., Varela, S., Jaramillo, C., Velez‐Juarbe, J., &
Silliman, B. R. (2016). Geographical distribution patterns of Carcharocles megalodon over time reveal clues about extinction mechanisms. Journal of Biogeography, 43(8), 1645-1655. https://doi.org/10.1111/jbi.12754
Riggs, S. R., Ames, D. V., Culver, S. J., & Mallinson, D. J. (2020). The battle for North Carolina’s coast: Evolutionary history, present crisis, and vision for the future.
Riggs, S. R., Cleary, W. J., & Snyder, S. W. (1995). Influence of inherited geologic framework on barrier shoreface morphology and dynamics. Marine Geology, 126(1-4), 213-234. https://doi.org/10.1016/0025-3227(95)00079-e
Schulte, P., Alegret, L., Arenillas, I., Arz, J., Barton, P. J., Brown, P. R., Bralower, T. J., Christeson, G. L., Claeys, P., & Willumsen, P. S. (2010). The Chicxulub Asteroid Impact and Mass Extinction at the Cretaceous-Paleogene Boundary. Science, 327(5970), 1214-1218. https://www.science.org/doi/10.1126/science.1177265
A breath breaks the air first — a soft exhale that sounds almost human — and then a dorsal fin lifts from the channel like a line drawn through moving water. The tide is falling. Gulls hover over the seam where current tightens. Fishermen pause mid-cast because everyone knows the rhythm: if the dolphins are working the edge, the fish are already gathering.
These encounters feel spontaneous, but they are not accidents. The dolphins that surface beside our piers, marsh creeks, and inlets are not anonymous travelers passing through. Many bottlenose dolphins show long-term site fidelity and structured community patterns in estuarine systems, returning to the same places across years (Urian et al., 2009; Wells, 2014). To live on this shoreline is to share space with minds moving just below the surface — residents of the tidal edge.
Who they are: a coastal population
The dolphins most frequently seen along Onslow County’s waters are common bottlenose dolphins (Tursiops truncatus), a species whose “coastal” lives can look very different from “offshore” lives. Across the western North Atlantic, genetic studies show fine-scale population structure that can separate dolphins using nearshore coastal waters from dolphins using inshore estuarine waters (Rosel et al., 2009). More broadly, integrative work continues to support meaningful coastal vs offshore divergence in the region (Costa et al., 2022).
In estuaries, photo-identification research (matching dorsal-fin markings) repeatedly shows that bottlenose dolphins can form discrete social communities with limited spatial overlap — a pattern consistent with long-term residency and local familiarity (Urian et al., 2009). In practical terms, the dolphin a child watches from a dock in spring may be seen again the following winter, and again the next year: not a rumor, but a biological possibility supported by long-term studies of resident dolphins elsewhere on the coast (Wells, 2014).
Photo-identification doesn’t always rely solely on human matching of fin shapes; new tools such as machine learning are being developed to improve accuracy in identifying individual dolphins and whales in the wild. For example, researchers in Hawaii are using advanced algorithms to distinguish individuals from large photo libraries of dorsal fins. As technology improves, methods like photo-ID only get more reliable — which means studies of habitat overlap and seasonal return become more precise over time.
An inside look at how scientists “read” dorsal fin shapes and markings to track the same dolphins over time.
Reading the geometry of the estuary
Dolphins do not simply occupy estuaries; they interpret them.
Tidal channels function as moving architecture. Falling tides compress fish schools toward narrowing exits. Sandbars redirect flow into faster seams. Marsh edges trap prey against shallow gradients. Dolphins exploit these features with precision, repeatedly targeting conditions that make prey capture more efficient (Barros & Wells, 1998; Torres & Read, 2009).
This is one reason dolphins so often appear where the water “looks alive” — at convergence lines, inlet throats, and channel bends. In Florida Bay, for example, foraging tactics are mapped onto habitat features that define where dolphins have spent their time, thus turning behavior into geography (Torres & Read, 2009). What seems like play from shore can be highly strategic predation.
Bottlenose dolphins breaching off Seaview Pier, N. Topsail Beach, North Carolina. The arc of the body and column spray reflect the mechanics of propulsion – force directed through the tail, momentum carried into the air. | Photo credit: Howard Crumpler Photography, 2026
Reader Question:
Why do dolphins seem more active on rainy or overcast days?
Weather, light, and the illusion of play
You may notice that dolphins seem especially active on overcast or rainy days — surfacing more frequently, breaching, or moving in tight arcs through wind-rippled water. It can look like preference, even mood. But dolphins are responding less to cloud cover than to what cloud cover does to the water.
When the sky darkens, baitfish don’t stay arranged the same way. They may bunch together or rise toward the surface. For a predator already working those upper layers, that shift can make hunting more efficient (Benoit-Bird & Au, 2003). Wind and rain can also stir the surface and cloud the water, changing who sees whom first (De Robertis et al., 2003).
There is also a perceptual component. Overcast skies reduce glare, making dorsal fins and splashes easier for human observers to detect. Wind-textured water highlights movement. What appears to be “more play” may sometimes be improved visibility — a reminder that observer experience and animal behavior are not always the same phenomenon.
In short, dolphins are responding to ecological conditions. The weather alters the water; the water alters the fish.
Two bottlenose dolphins break the surface beneath the gray horizon off Surf City, North Carolina. Overcast light and wind-roughened water can change how fish move – and how easily we notice the dolphins following them. | Photo credit: Johnny Provost, Jr., 2025
Communication and social intelligence
Bottlenose dolphins have been studied for decades not just because they are charismatic, but because their social lives depend on constant communication in a shifting, three-dimensional world. One of the strongest findings to emerge from that research is the existence of signature whistles — individually distinctive call types that function as learned identity signals, something very much like the individual name a dolphin goes by within its community (Janik & Sayigh, 2013).
Social learning runs just as deep. Some dolphin foraging habits spread from one animal to another rather than through genetics — passed along socially, a rare pattern among nonhuman species (Krützen et al., 2005). Mothers and calves stay together for years, giving calves time to learn not just how to hunt, but where — which channels to follow, which bends of water hold fish (Wells, 2014).
In some populations elsewhere in the world, dolphins even use tools — carrying marine sponges on their rostrums while foraging or trapping fish inside empty shells — behaviors that are socially learned and culturally transmitted (Krützen et al., 2005).
That learning shapes how dolphins fit into the estuary. In many tidal systems they sit near the top of the local food web, influencing the fish communities beneath them. Yet beyond those protected waters, they are not beyond risk. Large sharks prey on dolphins, placing them within a broader coastal hierarchy where even predators can become prey (Heithaus, 2001). The role shifts with scale. The ecology remains layered.
Two bottlenose dolphins surfacing together off Seaview Pier, N. Topsail Beach, North Carolina. Close positioning and timing are hallmarks of the complex social bonds that define dolphin societies. | Photo credit: Howard Crumpler Photography, 2026
Dolphins are not guardians
Popular culture has assigned dolphins a role they never chose: protector. People repeat a comforting shoreline myth — “If you’re scared of sharks, find the dolphins; they’ll protect you.” But that story is not grounded in how dolphins behave in the wild.
Bottlenose dolphins are powerful predators. They compete, establish dominance hierarchies, and can deliver forceful blows when defending calves or asserting space. Dolphin–shark interactions occur, but they are not “rescue missions” staged for humans; they are ecological encounters shaped by risk, competition, and opportunity (Heithaus, 2001).
Wild dolphins are also capable of injuring people. Research examining human–dolphin interactions show that close approaches — and especially feeding wild dolphins — increase the likelihood of risky contact and harmful outcomes for both dolphins and people (Cunningham-Smith et al., 2006; Vail, 2016). Over time, those interactions leave visible consequences. Long-term data from Sarasota Bay show that dolphins who have learned to associate people with food are more likely to carry injuries linked to boats and fishing gear (Christiansen et al., 2016).
The danger is not that dolphins are “evil.” The danger is assuming they share human intentions.
Swimming near a pod does not create a protective shield. Dolphins are not lifeguards. They are wild animals navigating their own priorities in a shared environment. Respecting that boundary is what allows coexistence.
A bottlenose dolphin pursuing prey near a recreational vessel in a waterway in Surf City, North Carolina. Foraging behavior can bring dolphins into close proximity with boats – not as companions, but as active predators focused on fish. | Video credit: Cynthia Dirosse, 2024
Winter dolphins
A persistent assumption is that dolphins vanish when the water cools. In reality, seasonal distribution can be more nuanced — changing with prey, temperature, and coastal movement patterns rather than following a simple on/off presence.
Along the mid-Atlantic coast, research shows that bottlenose dolphins shift their movements with the seasons, appearing in different areas at different times of year (Torres et al., 2005). Studies focused on estuarine dolphins in southern North Carolina document similar seasonal patterns closer to home (Silva et al., 2020). From shore, those changes can look like disappearance. But winter quiet does not always mean absence. It may simply mean dolphins are working deeper channels or less visible pathways beyond the easy reach of our eyes.
The estuary in winter is quieter, but not empty.
Dorsal fins in winter light off Surf City, North Carolina. Dolphins may appear less active this time of year, but changes in light, water depth, and travel corridors often influence what we notice from shore. | Photo credit: Surf City Parks, Recreation, and Tourism, 2017
Living beside them
Living near dolphins is a privilege — and it places us within the same waters they navigate. Vessel traffic, fishing gear, and repeated close approaches can shape the lives of animals that live for decades and raise calves slowly (Wells, 2014). Studies of dolphins that have been fed or closely approached by people show that these interactions can shift behavior, making dolphins more likely to approach boats and increasing the risk of injury and conflict (Vail, 2016). Distance, in that sense, preserves the patterns people come to watch.
The presence of dolphins is not guaranteed. It is a sign that the system still functions — prey, water quality, shoreline structure, and the complex social knowledge dolphins carry from year to year. As long-lived predators near the top of the food web, they are indicator species, reflecting the condition of the waters they inhabit — estuary, inlet, and nearshore coast alike.
And so when a dorsal fin rises beyond the channel markers, it means more than a moment of spectacle. It means the currents are still working, the fish are still moving, and the layered relationships that shape this shoreline are still holding.
There is always more to learn about dolphins than fits in a single post. For those who’d like to go further, this episode of the All Creatures Podcast offers a thoughtful exploration of their biology and behavior.
References
Barros, N. B., Wells, R. S., & Barros, N. B. (1998). Prey and feeding patterns of resident bottlenose dolphins (Tursiops truncatus) in Sarasota Bay, Florida. Journal of Mammalogy, 79(3), 1045. https://doi.org/10.2307/1383114
Benoit-Bird, K. J., & Au, W. W. (2003). Prey dynamics affect foraging by a pelagic predator (Stenella longirostris) over a range of spatial and temporal scales. Behavioral Ecology and Sociobiology, 53(6), 364-373. https://doi.org/10.1007/s00265-003-0585-4
Christiansen, F., McHugh, K. A., Bejder, L., Siegal, E. M., Lusseau, D., McCabe, E. B., Lovewell, G., & Wells, R. S. (2016). Food provisioning increases the risk of injury in a long-lived marine top predator. Royal Society Open Science, 3(12), 160560. https://doi.org/10.1098/rsos.160560
Costa, A. P., Mcfee, W., Wilcox, L. A., Archer, F. I., & Rosel, P. E. (2022). The common bottlenose dolphin (Tursiops truncatus) ecotypes of the western North Atlantic revisited: An integrative taxonomic investigation supports the presence of distinct species. Zoological Journal of the Linnean Society, 196(4), 1608-1636. https://doi.org/10.1093/zoolinnean/zlac025
Cunningham-Smith, P., Colbert, D. E., Wells, R. S., & Speakman, T. (2006). Evaluation of human interactions with a provisioned wild bottlenose dolphin (<I>Tursiops truncatus</I>) near Sarasota Bay, Florida, and efforts to curtail the interactions. Aquatic Mammals, 32(3), 346-356. https://doi.org/10.1578/am.32.3.2006.346
De Robertis, A., Ryer, C. H., Veloza, A., & Brodeur, R. D. (2003). Differential effects of turbidity on prey consumption of piscivorous and planktivorous fish. Canadian Journal of Fisheries and Aquatic Sciences, 60(12), 1517-1526. https://doi.org/10.1139/f03-123
Heithaus, M. R. (2001). Shark attacks on bottlenose dolphins (TURSIOPS ADUNCUS) in Shark Bay, Western Australia: Attack rate, bite scar frequencies, and attack seasonality. Marine Mammal Science, 17(3), 526-539. https://doi.org/10.1111/j.1748-7692.2001.tb01002.x
Janik, V. M., & Sayigh, L. S. (2013). Communication in bottlenose dolphins: 50 years of signature whistle research. Journal of Comparative Physiology A, 199(6), 479-489. https://doi.org/10.1007/s00359-013-0817-7
Krützen, M., Mann, J., Heithaus, M. R., Connor, R. C., Bejder, L., & Sherwin, W. B. (2005). Cultural transmission of tool use in bottlenose dolphins. Proceedings of the National Academy of Sciences, 102(25), 8939-8943. https://doi.org/10.1073/pnas.0500232102
Rosel, P. E., Hansen, L., & Hohn, A. A. (2009). Restricted dispersal in a continuously distributed marine species: Common bottlenose dolphinsTursiops truncatusin coastal waters of the western North Atlantic. Molecular Ecology, 18(24), 5030-5045. https://doi.org/10.1111/j.1365-294x.2009.04413.x
Silva, D. (2020). Abundance and seasonal distribution of the southern North Carolina estuarine system stock (USA) of common bottlenose dolphins (Tursiops truncatus). IWC Journal of Cetacean Research and Management, 21(1), 33-43. https://doi.org/10.47536/jcrm.v21i1.175
Torres, L. G., McLellan, W. A., Meagher, E., & Pabst, D. A. (2023). Seasonal distribution and relative abundance of bottlenose dolphins, Tursiops truncatus, along the US Mid-Atlantic coast. J. Cetacean Res. Manage, 7(2), 153-161. https://doi.org/10.47536/jcrm.v7i2.748
Torres, L. G., & Read, A. J. (2009). Where to catch a fish? The influence of foraging tactics on the ecology of bottlenose dolphins (Tursiops truncatus) in Florida Bay, Florida. Marine Mammal Science, 25(4), 797-815. https://doi.org/10.1111/j.1748-7692.2009.00297.x
Urian, K. W., Hofmann, S., Wells, R. S., & Read, A. J. (2009). Fine‐scale population structure of bottlenose dolphins (Tursiops truncatus) in Tampa Bay, Florida. Marine Mammal Science, 25(3), 619-638. https://doi.org/10.1111/j.1748-7692.2009.00284.x
Vail, C. S. (2016). An overview of increasing incidents of bottlenose dolphin harassment in the Gulf of Mexico and possible solutions. Frontiers in Marine Science, 3. https://doi.org/10.3389/fmars.2016.00110
Wells, R. S. (2013). Social structure and life history of bottlenose dolphins near Sarasota Bay, Florida: Insights from four decades and five generations. Primatology Monographs, 149-172.
At low tide in winter the creek mouths behind Topsail Island widen into ground that is usually concealed, and the exposed marsh does not appear emptied so much as translated into another state where water has thinned into channels narrow enough to reveal the structure it normally masks. The flats emerge as a textured plane stitched by the remains of Spartina alterniflora, each stem cluster surrounded by a faint collar of darker mud where drainage lags by seconds, and the surface separates into alternating bands that hold or soften depending on how recently porewater escaped. This firmness reflects sediment consolidation, the gradual compression of mud as water drains between tides, tightening elevated shelves first and leaving adjacent troughs saturated, so the exposed ground becomes a map of load-bearing ridges that anticipates where larger animals will move once the marsh opens (Christiansen et al., 2000; Morris et al., 2002).
Close to the surface, winter resolves into finer evidence that the marsh is neither dormant nor still. Fiddler crab chimneys crumble into damp grains that expose darker sediment beneath a thin crust, while hoofprints from the previous tide hold shallow mirrors rimmed with frost where a faint olive sheen gathers as diatoms trap warmth and moisture (Underwood & Kromkamp, 1999). Beside the prints, spirals of fine sediment rise like coiled handwriting, polychaete casts lifted from below and dried into granular ridges that record upward movement from buried layers. Every centimeter of mud registers exchange between subsurface metabolism and cold air, and the exposed flats behave less like the absence of water than a temporary reorganization of it, one that prepares a surface already structured for the next set of crossings.
Clusters of crab burrow openings mark the marsh surface, each hole a vertical conduit linking oxygen, water, and nutrients to the sediment below. | Photo credit: M. Mitchell, 2026
Where winter concentrates energy
The winter low tide exposes more than terrain, because the withdrawal of water aligns accessibility with abundance in a way that concentrates food at the surface for a brief interval. Spartina rhizomes lie just beneath the crust, their pale ends visible where deer have bitten through the mud, and detached stems gather in wrack lines where microbial films soften fibrous blades into digestible pulp. Small bivalves remain gaping in shallow pools where temperature lingers above the surrounding flats, and worm casts cluster where organic matter has settled densely enough to support continuous feeding below. This alignment functions as a resource pulse, a moment when energy stored in buried plant tissue and invertebrate biomass becomes reachable simultaneously.
Wrack concentrated by winter tides stores organic energy in dense bands, drawing shorebirds to feed where nutrients accumulate along the marsh edge. | Photo credit: American Birding Association
Deer enter the marsh along consolidated ridges that hold their weight, yet the crossings do not run straight through these zones of exposure but instead loop and return around feeding sites where sediment has been churned darker than its surroundings. The mud at these points holds fragments of torn rhizomes pressed into its surface and shredded plant fibers mixed into the crust, while overlapping tracks form shallow basins that later fill with water and preserve the geometry of the feeding circuit. Raccoon prints braid across the same lines, Canada goose droppings mark cropped stems, and dunlin and greater yellowlegs settle repeatedly where the surface softens under pressure, their bills puncturing the crust in arcs that echo the paths carved by hooves. Exposure redistributes energy upward, and movement gathers along the same ridges that consolidation established, tying feeding to structure without separating the two processes.
The skin that reforms
Cross-section of marsh sediment showing deposition, erosion, and consolidation, the shifting layers that form and reform the exposed winter surface. | Graphic credit: G. S. Sylvain, 2011
Between exposures, slack water leaves a thin veneer that dries into a continuous surface film through sediment sealing, a layer fine enough to slow the exchange of gases between air and mud (Christiansen et al., 2000). When intact, the flats dull into a flexible sheet that bends faintly under weight, and breaking it releases a muted sulfur odor that signals redox cycling, the shift between oxygenated and oxygen-poor states driven by microbial respiration in buried sediment (Howarth & Teal, 1979; Mendelssohn et al., 1981). Color reveals the chemistry more reliably than smell. Black veins branch through exposed mud where iron binds sulfide, while pale halos surround Spartina roots where oxygen leaks downward along living tissues.
Each footprint becomes an aperture in this membrane, allowing oxygen to enter and reduced compounds to rise, so the breach brightens temporarily before darkening again as metabolism rebalances. Feeding animals convert chemical gradients into visible patterns, and the flats accumulate a shifting mosaic of sealed and reopened zones that migrate with every tide, ensuring that the next exposure inherits the chemical memory of the previous one.
Tracks fracture the sealed winter crust, revealing darker sediment where oxygen re-enters and the surface begins to reform. | Photo Credit: M. Gold, 2023
The ground below the ground
Beneath the crust, the sediment continues to reorganize through bioturbation, the mixing of mud by infaunal animals whose activity does not cease with falling temperature. Polychaete worms thread galleries through the upper layers, lifting sediment to the surface in tight spirals while their burrows act as ventilation shafts through burrow ventilation, drawing oxygen downward and leaking reduced porewater upward (Kristensen, 2000; Aller, 1982). Small bivalves pump water through siphons that leave paired pinholes scattered across the flats, and amphipods graze biofilms coating the worm casts, linking subsurface feeding to surface texture.
Each round of burrowing lifts buried debris and nutrients toward the surface, making crab tunnels pathways that continually rebuild the marsh from below. | Graphic credit: Wang et al., 2010
Where deer cross and feed, hooves collapse some tunnels while sealing others, producing prints that darken unevenly because subsurface architecture differs from step to step. The feeding circuits therefore overlay hidden engineering that maintains permeability and redistributes nutrients, ensuring that exposure, grazing, and burrowing operate as one continuous process rather than as isolated events separated by layers of mud.
Smell in shallow water
Disturbed sediment releases dissolved compounds that spread through shallow pools as porewater plumes, chemical gradients that extend beyond the visible cloud of suspended mud. Killifish and juvenile mullet navigate these gradients through chemoreception, keeping their snouts close to the surface while pivoting toward intensifying scent (Kneib, 1997; Kristensen, 2000). Their feeding loosens additional sediment and amplifies the plume before particles settle again, creating a moving field of chemical information that overlaps with the physical contours of the flats.
What appears from above as a brief swirl becomes a signal that attracts birds, and dunlin and yellowlegs converge on fresh pits where worms remain exposed. Each crater fills with water and darkens as sulfide seeps upward, and feeding layers stack in sequence so that invertebrate disturbance leads to fish excavation, which leads to avian probing, all anchored to the same exposure that first drew deer into the marsh. Leaning close reveals faint popping as methane and carbon dioxide escape through gas ebullition, ticking upward from saturated sediment while animals feed across the surface. The marsh ventilates audibly, and the sound marks exchange continuing beneath apparent stillness.
As the tide withdraws, exposed mud concentrates scent and invertebrates near the surface, guiding shorebirds to feeding zones written into the sediment. | Photo credit: Ron Watts
Memory in the surface
Winter tides and storms deposit sediment that raises the marsh through vertical accretion, stacking particles in increments small enough to disappear into the surface unless read over time (Morris et al., 2002). Hurricane overwash leaves thin sand sheets that redirect drainage for months, oyster clusters trap suspended grains in their lee (Newell et al., 2005), and worm burrows stabilize some deposits while loosening others (Kirwan & Megonigal, 2013). Feeding compresses ridges and excavation softens troughs, embedding each disturbance into the next layer so that the flats carry a structural memory of their own use.
Returning after weeks reveals crossings shifted, wrack lines buried, and worm casts clustered in new zones, evidence that the marsh does not reset between exposures but accumulates the imprint of repeated winter engineering.
Winters that change
Warmer temperatures extend microbial activity through temperature-driven metabolic acceleration, thinning the interval between sealing and decay and allowing chemical gradients to persist longer at the surface (Bridgham et al., 2006). Rising water levels narrow exposure windows, stronger storms redistribute sediment in thicker pulses, and shifting coastal currents alter nutrient delivery and larval supply, influencing which species occupy the winter flats (Kirwan & Megonigal, 2013). The marsh continues to open, yet the rhythm of exposure recalibrates, and feeding circuits migrate toward higher shelves where consolidation still holds.
Chemical plumes stretch farther in warmer water, grazing concentrates into narrower bands, and the same negotiations between structure and feeding repeat under altered timing, ensuring that winter engineering continues without preserving its previous schedule.
Seasonal temperature outlook showing shifting winter probabilities across the southeastern United States, a regional signal that filters down to marsh-level processes. | NOAA – National Weather Service, 2026
The surface in motion
The creek mouth appears quiet until attention lowers to the scale of sediment. Frost melts along print rims before surrounding crust warms, gas ticks upward through worm tubes, fish pits refill, and diatoms bloom where warmth collects. Each tide writes another layer into a system held in dynamic equilibrium, continuous adjustment that maintains form while never remaining fixed (Morris et al., 2002). Exposure leads to feeding, feeding reshapes structure, and structure governs the next exposure as the marsh opens again.
The same ridges that hold a deer’s weight will soften again when the tide returns, and the feeding circuits traced across them will dissolve into channels that redistribute the next layer of sediment. Worm burrows will reopen where hooves sealed them, chemical plumes will reassemble in newly flooded pools, and the surface will carry forward the imprint of this exposure into the next one. Winter does not suspend the marsh. It recalculates it at a slower tempo, redistributing energy across the same structures that will support spring growth and summer density, so that even in the coldest intervals the creek mouth continues its quiet accounting of exchange, preparing another surface that will open and be read again.
Deer cross the marsh to reach winter feeding exposed by the tide, moving along corridors that appear only when the surface opens. | Photo credit: L. W. Hamilton, 2025
References
Aller, R. C. (1982). The effects of Macrobenthos on chemical properties of marine sediment and overlying water. Topics in Geobiology, 53-102. https://doi.org/10.1007/978-1-4757-1317-6_2
Christiansen, T., Wiberg, P., & Milligan, T. (2000). Flow and sediment transport on a tidal salt marsh surface. Estuarine, Coastal and Shelf Science, 50(3), 315-331. https://doi.org/10.1006/ecss.2000.0548
Howarth, R. W., & Teal, J. M. (1979). Sulfate reduction in a New England salt marsh1. Limnology and Oceanography, 24(6), 999-1013. https://doi.org/10.4319/lo.1979.24.6.0999
Kirwan, M. L., & Megonigal, J. P. (2013). Tidal wetland stability in the face of human impacts and sea-level rise. Nature, 504(7478), 53-60. https://doi.org/10.1038/nature12856
Kneib, R. T. (1997). The role of tidal marshes in the ecology of estuarine nekton. Oceanography And Marine Biology, 35(35), 159-216. https://doi.org/10.1201/b12590-5
Kristensen, E. (2000). Organic matter diagenesis at the oxic/anoxic interface in coastal marine sediments, with emphasis on the role of burrowing animals. Hydrobiologia, 426(1), 1-24. https://doi.org/10.1023/a:1003980226194
Mendelssohn, I. A., McKee, K. L., & Patrick, W. H. (1981). Oxygen deficiency in Spartina alterniflora roots: Metabolic adaptation to anoxia. Science, 214(4519), 439-441. https://doi.org/10.1126/science.214.4519.439
Morris, J. T., Sundareshwar, P. V., Nietch, C. T., Kjerfve, B., & Cahoon, D. R. (2002). Responses of coastal wetlands to rising sea level. Ecology, 83(10), 2869. https://doi.org/10.2307/3072022
Newell, R. I., Fisher, T. R., Holyoke, R. R., & Cornwell, J. C. (2005). Influence of eastern oysters on nitrogen and phosphorus regeneration in Chesapeake Bay, USA. NATO Science Series IV: Earth and Environmental Series, 282, 93-120. https://doi.org/10.1007/1-4020-3030-4_6
Underwood, G., & Kromkamp, J. (1999). Primary production by phytoplankton and Microphytobenthos in estuaries. Advances in Ecological Research, 29, 93-153. https://doi.org/10.1016/s0065-2504(08)60192-0
A microscopic epic of drift, decision, and devotion
On a winter walk along a pier in Surf City, the boards are bleached pale by sun and salt. Wind threads through the pilings. Gulls cry over gray water. At your feet, on a beam that has known decades of tides, something clings.
It is no bigger than a fingernail—chalky white, ridged like a tiny volcano. Along this coast, it is often an ivory barnacle—Amphibalanus eburneus—one of the small architects that quietly carpet pilings, docks, and seawalls from Topsail Sound to the Cape Fear. You could scrape it away with the edge of a shell. You probably have, absentmindedly, a hundred times.
But this barnacle is not debris. It is a biography written in calcium.
It began as a drifting dot—an invisible life in a moving sea. It crossed currents. It tasted the chemistry of places. And then, once, it chose.
The choice was final.
Barnacles are among the few animals on Earth that get exactly one chance to decide where they will live. No revisions. No migrations. No second homes. The place where a barnacle settles becomes the place where it will eat, grow, reproduce, and die. Its entire life collapses into a single coordinate on the map of the shore.
To understand a barnacle is to understand what it means to commit.
Ivory barnacles cling to a rock | Photo credit: Ken-ichi Ueda
Drift
A barnacle’s life begins in motion.
After fertilization, barnacle embryos hatch into nauplius larvae—tiny, triangular forms equipped with beating appendages and a simple eye (Anderson, 1994). They rise into the plankton, where they may drift for days to weeks, feeding and growing as tides and currents carry them outward (Chen et al., 2014).
The first larval stage of a barnacle, called a nauplius, is free-swimming and distinguished by a set of “horns.” | Photo credit: Robert Bachand
They are not aimless. Even at this scale, nauplii respond to light, salinity, and gravity. They migrate vertically through the water column, riding layers of current like conveyor belts. Their world is vast and borderless—and lethal.
Most barnacles die here.
Nauplii are eaten by copepods, jellyfish, fish larvae, and filter-feeding invertebrates. Each pulse of water is a gauntlet. Survival depends on number: millions released so that a few may reach shore.
After several molts, the nauplius enters its final larval form: the cyprid.
A late larval barnacle stage, the cyprid, has a bivalved shell of chitin and glands in its first antennae that are used to cement itself permanently to a hard substrate. | Photo credit: Robert Bachand
This is no longer a feeding animal. It is a vessel of stored energy, built for a single task—finding a place to live (Aldred & Clare, 2008).
The cyprid does not eat.
A clock begins.
Much of what we know about this hidden stage comes from decades of work on a close coastal relative, the striped barnacle – Amphibalanus amphitrite—a warm-water barnacle that clings to pilings and boat hulls worldwide, and whose larvae have become a window into how barnacles read the sea.
The striped barnacle (Amphibalanus amphitrite) is a globally distributed, non-native barnacle species that can spread via biofouling. In North Carolina waters it may occur outside its historical native range, but it isn’t widely recognized as a documented invasive species causing major ecological disruption. | Photo Credit: South Australia Marine Lab
The Narrow Window
Now the barnacle is no longer drifting blindly. It swims with intent. The cyprid probes surfaces with specialized antennules, “tasting” the chemistry of rock, wood, shell, and steel. It detects microbial biofilms—thin living skins that signal a surface has been stable long enough to support life (Qian et al., 2007). It senses the presence of other barnacles. It avoids surfaces that feel wrong.
This sensory world evolved in seas that were chemically simpler.
Today, cyprids swim through waters laced with heavy metals, hydrocarbons, microplastics, antifouling compounds, and nutrient-driven microbial shifts. These pollutants alter biofilms, mask settlement cues, and interfere with larval sensory systems. What once read clearly as “home” now arrives as static.
In degraded waters, cyprids often hesitate. They probe and retreat. They circle without committing.
But the clock does not pause.
Depending on species and temperature, a cyprid has only days to a few weeks before its stored energy is exhausted (Aldred & Clare, 2008). Each hour of searching burns fuel. When reserves fall too low, three futures unfold.
Some larvae simply die in the plankton and sink.
Some make a desperate choice—cementing themselves to marginal or unstable surfaces.
Others respond to distorted cues and settle where survival is unlikely.
This is not a failure of instinct. It is a mismatch between ancient sensory logic and a changed sea.
Long before we notice a shoreline growing quieter, its future has already thinned in the plankton.
When the answer is yes, the barnacle performs one of the most irreversible acts in the animal kingdom.
It flips upside down.
Using its antennules, the cyprid secretes a permanent biological cement and glues its head to the surface (Kamino, 2016). This adhesive—among the strongest natural glues known—binds underwater to stone, metal, and polymer. Once cured, it cannot be undone.
There is no “testing.” No trial period.
This is the end of motion.
Within hours, the cyprid undergoes a radical metamorphosis. Its eyes degenerate. Its swimming limbs are restructured into feathery feeding appendages called cirri. Its body reorganizes around a new axis—rooted instead of free (Høeg & Møller, 2006).
The barnacle becomes architecture.
Many do not survive even this. Newly settled juveniles are grazed by small fish and invertebrates. Waves scrape them away before cement fully cures. The shoreline is littered with choices that did not last.
Those that remain begin to build something larger than themselves.
A Life Built Around the Tide
Most animals grow by addition. Barnacles grow by reinvention.
Shell plates rise around soft tissue, forming a fortress against wave impact, desiccation, and predation. Inside, muscles and organs reorganize to support a life of rhythmic feeding.
When submerged, the barnacle opens its opercular plates and unfurls its cirri—six pairs of jointed limbs that sweep the water in steady arcs. Each beat captures phytoplankton, detritus, and microcrustaceans (Southward, 2008).
Metabolism slows. Heat and salt concentrate. Time folds inward. Some intertidal barnacles endure body temperatures exceeding 40°C (104°F) and prolonged oxygen deprivation (Harley, 2008). They wait for the sea to return.
Each tide is both a threat and nourishment.
Anatomy of a barnacle. | Photo Credit: AnimalFact.com
Time in Shell
Barnacles record time the way trees do.
Their shells grow in increments, forming visible growth bands that reflect seasonal cycles and environmental stress (Crisp, 1989). Storms leave signatures. Cold winters slow deposition. Productive summers thicken walls.
A barnacle on a piling may live five, ten, even twenty years (Southward, 2008). It will experience thousands of tides, hundreds of storms, and uncountable shifts in salinity and temperature—without ever moving.
Where foraminifera archive ancient seas in sediment, barnacles archive living shorelines in calcium.
Barnacles occupy one of the most punishing habitats on Earth: the intertidal zone.
Here, organisms must withstand:
Wave forces exceeding hurricane winds
Repeated drying and rehydration
Rapid temperature swings
Salinity changes from rain and evaporation
Intense ultraviolet exposure
Few animals can survive here. Barnacles not only survive—they structure the place.
Every barnacle on this shore is the consequence of a single larval decision made weeks earlier in open water.
They stabilize surfaces. They retain moisture. They create crevices for algae, worms, snails, and juvenile crustaceans. They shape temperature gradients and water flow. They turn bare rock into habitat.
When settlement falters—when larvae cannot read the shore or run out of time—the architecture of the coast changes.
Bare rock expands. Algal communities shift. Grazers lose shelter. Predators lose prey. The intertidal simplifies.
A piling with fewer barnacles is not merely cleaner. It is quieter. Biologically poorer and less layered.
The Lesson in Shell
Return now to that single barnacle on the pier.
It has no eyes. It has never seen the ocean. It will never know the gull overhead or the human who pauses above it. And yet it has shaped its entire existence around this exact sliver of coast.
It did not choose perfectly.
Some barnacles settle too high and starve. Some attach where sand scours them away. Some cement themselves beside competitors that outgrow and smother them.
There is no guarantee.
Only the act of choosing.
In a world that prizes movement, flexibility, and endless revision, the barnacle offers a quieter philosophy:
At some point, life must become a place.
To belong is not to drift forever. It is to accept exposure. To endure storms. To open when the tide allows. To grow, layer by layer, into the shape of your ground.
Every barnacle on this coast is a monument to a single irreversible decision.
And the sea is full of them.
Bay barnacle, Amphibalanus improvisus, on a rock in the New River | Photo credit: Alina Michele, iNaturalist, 2022
References
Aldred, N., & Clare, A. S. (2008). The adhesive strategies of cyprids and development of barnacle-resistant marine coatings. Biofouling, 24(5), 351-363. https://doi.org/10.1080/08927010802256117
Anderson, D. T. (1994). Barnacles: Structure, function, development and evolution (1st ed.). Springer Dordrecht.
Chen, Z., Zhang, H., Wang, H., Matsumura, K., Wong, Y. H., Ravasi, T., & Qian, P. (2014). Quantitative Proteomics study of larval settlement in the barnacle balanus Amphitrite. PLoS ONE, 9(2), e88744. https://doi.org/10.1371/journal.pone.0088744
Crisp, D. J. (1989). Tidally deposited bands in shells of barnacles and molluscs. Origin, Evolution, and Modern Aspects of Biomineralization in Plants and Animals, 103-124. https://doi.org/10.1007/978-1-4757-6114-6_8
Harley, C. D. (2008). Tidal dynamics, topographic orientation, and temperature-mediated mass mortalities on rocky shores. Marine Ecology Progress Series, 371, 37-46. https://doi.org/10.3354/meps07711
Høeg, J. T., & Møller, O. S. (2006). When similar beginnings lead to different ends: Constraints and diversity in cirripede larval development. Invertebrate Reproduction & Development, 49(3), 125-142. https://doi.org/10.1080/07924259.2006.9652204
Qian, P., Lau, S. C., Dahms, H., Dobretsov, S., & Harder, T. (2007). Marine Biofilms as mediators of colonization by marine Macroorganisms: Implications for antifouling and aquaculture. Marine Biotechnology, 9(4), 399-410. https://doi.org/10.1007/s10126-007-9001-9
Southward, A. J. (2008). Barnacles: Keys and notes for the identification of British species. Field Studies Council. Yu, S., & Chan, B. K. (2020). Intergenerational microplastics impact the intertidal barnacle Amphibalanus Amphitrite during the planktonic larval and benthic adult stages. Environmental Pollution, 267, 115560. https://doi.org/10.1016/j.envpol.2020.115560
How seagrasses and marsh grasses—and the animals within them—build the marshes of Onslow County
In Onslow County’s estuarine marshes, the best time to understand how the landscape works is when the water pulls back. As tides drain from creeks and shallow flats, patterns begin to emerge—where water lingers, where it moves easily, and where it hesitates. These patterns are not random. They reflect the combined influence of plants, animals, and sediments continually reshaping the boundary between land and sea.
Like the microscopic shells of foraminifera preserved in sediment, marsh and seagrass communities record environmental conditions. But unlike the past locked in mud, these systems are alive, constantly negotiated by plants, grazers, predators, and microbes.
From permanently submerged seagrass beds to the highest marsh edge, each elevation zone in Onslow County is maintained not just by vegetation, but by species that actively regulate growth, chemistry, and water flow.
Subtidal shallows: seagrass beds maintained by grazers
In the shallow, light-penetrated waters of the New River Estuary and protected soundside areas, seagrass beds form underwater meadows that stabilize sediments and provide nursery habitat for fish and invertebrates. Species present or expected in Onslow County waters include eelgrass (Zostera marina), shoalgrass (Halodule wrightii), and widgeongrass (Ruppia maritima) (Mallin, 2000; Orth, 1984).
Seagrass blades rapidly accumulate epiphytic algae and microbial films. Without constant grazing, this layer can block light and suppress photosynthesis. Amphipods, isopods, and small gastropods act as continuous maintenance crews, grazing epiphytes and preventing them from overwhelming the plants themselves (Orth & van Montfrans, 1984; Valentine & Duffy, 2006).
Experimental studies show that when these grazers are removed, seagrass condition declines even under favorable light conditions, demonstrating that plant survival depends as much on animal activity as on physical environment (Duffy et al., 2015). Beneath the canopy, burrowing worms and bivalves recycle nutrients and oxygenate sediments, preventing organic matter from accumulating around roots (Orth, 1984).
In this zone, seagrass persists because grazers keep blades clean and sediments breathable—a cooperative system built on constant biological upkeep.
Gammarus mucronatus, a common amphipod grazer on eelgrass | Photo credit: E. A. Lazo-Wasem, Yale Peabody Museum, 2013.
The low marsh edge: cordgrass shaped by snails and crabs
At the daily-flooded edge of the marsh, smooth cordgrass (Spartina alterniflora) dominates. This narrow fringe marks the boundary between open water and marsh interior, where erosion pressure is highest and stability matters most.
Left: Healthy smooth cordgrass (Spartina alterniflora) line the estuary edge in Surf City, NC. | Photo credit: A. Mitchell, 2022.Right: Salt marsh die-off from grazing stress by marsh periwinkle snails and reduced predation by crabs, such as blue crabs, can create bare mudflats. | Photo credit: By Esuglia at English Wikipedia, CC BY-SA 3.0, https://commons.wikimedia.org/w/index.php?curid=65794096
Cordgrass growth here is tightly regulated by the marsh periwinkle snail (Littoraria irrorata). These snails climb grass stems to avoid inundation and graze directly on living tissue, often intensifying damage by facilitating fungal infection. At high densities, periwinkle grazing can dramatically reduce cordgrass height and biomass, effectively mowing the marsh edge (Silliman & Zieman, 2001).
Marsh periwinkle snails (Littoraria irrorata) are a common sight on cordgrass (Spartina alterniflora) in North Carolina | Photo credit: North Carolina Aquarium at Roanoke Island, 2018.
Unchecked grazing can destabilize the marsh platform—but periwinkles themselves are regulated by crabs, including blue crabs (Callinectes sapidus),fiddler crabs (Genus Uca), purple marsh crabs (Sesarma reticulatum), hermit crabs and other burrowing species. Crabs prey on snails, limiting grazing pressure and indirectly protecting cordgrass (Silliman et al., 2005).
Crabs also function as ecosystem engineers. Their burrows aerate sediments, relieve sulfide stress around plant roots, and improve tidal water movement through compacted soils (Bertness, 1985; Thomas & Blum, 2010). Where crabs are abundant, cordgrass grows taller and denser; where they are lost, marsh die-off can occur rapidly.
This zone persists through a trophic cascade: grass builds land, snails limit grass, and crabs keep the system in balance.
Mid-marsh: mussels and detritus processors reinforce the platform
Just upslope, where flooding becomes less frequent, plant communities shift toward mixtures that often include saltmeadow cordgrass (Spartina patens). Here, the ribbed mussel (Geukensia demissa) emerges as a key stabilizing force.
Saltmeadow cordgrass (Spartina patens) is an important marsh stabilizer that has higher productivity when it grows near ribbed mussel aggregations | Photo credit: Kristie Gianopulos
Ribbed mussels form dense clusters at the base of marsh vegetation, binding sediments with byssal threads and physically reinforcing marsh soils against erosion (Bertness, 1984). As filter-feeders, they concentrate nutrients by removing organic matter from tidal waters and depositing nitrogen-rich biodeposits directly into marsh sediments (Jordan & Valiela, 1982).
Ribbed mussels (Geukensia demissa) at the base of marsh vegetation | Photo credit: R. Bachand
Grasses growing near mussel aggregations exhibit higher productivity than those without mussels, demonstrating a strong facilitative relationship between animals and plants (Bertness, 1984). As vegetation senesces, detritivorous worms, insects, and microbial decomposers break down dead plant material, converting standing biomass into detritus that fuels food webs throughout the estuary (Mann, 1988).
The mid-marsh functions as a processing zone, reinforcing marsh structure while converting plant matter into usable energy.
High marsh: microbes that manage chemical stress
In the high marsh, dominated by black needlerush (Juncus roemerianus) and saltmeadow cordgrass (Spartina patens), flooding is limited to spring tides and storms. Prolonged exposure to air creates harsh soil conditions, including elevated salinity and sulfide accumulation.
Here, microbial communities play a central role. Sulfate-reducing and sulfur-oxidizing bacteria regulate sulfide concentrations that would otherwise become toxic to plant roots, while microbial decomposition controls nutrient availability under fluctuating oxygen conditions (Howarth & Giblin, 1983).
Beneath the marsh surface, soil microbes regulate decomposition, carbon exchange, and chemical stress. Changes in salinity and flooding reshape microbial communities, influencing how marsh soils process organic matter and support vegetation across tidal elevations. | Image credit: Zhang et al., 2023.
Small soil invertebrates maintain pore spaces that allow brief pulses of oxygenated water to penetrate during flooding. Unlike the visibly engineered low marsh, the high marsh is stabilized largely through biogeochemical regulation rather than grazing or predation.
This zone endures because microbes quietly buffer plants against chemical extremes.
From microbes in the soil to grasses at the surface, biological interactions drive marsh formation. Microbial processes govern decomposition and organic matter buildup, helping determine whether marsh platforms gain elevation, remain stable, or collapse | Image credit: Abbot, Quirk & Fultz, 2022.
The marsh–upland transition: keeping the boundary intact
At the uppermost margin of the marsh, tidal influence becomes intermittent and environmental stress shifts from salinity to erosion and freshwater input. Burrowing invertebrates increase soil permeability, allowing stormwater and tidal surges to infiltrate rather than scour the surface (Thomas & Blum, 2010).
A profile illustration . depicting the recommended transition of plant types from the edge of the salt marsh to the upland buffer. | Image credit: Massachusetts Office of Coastal Zone Management
Vegetation root networks stabilize soils exposed to drying and wave action, while animal burrows act as pressure-release pathways during extreme events. When these biological processes are disrupted—by shoreline hardening or vegetation removal—the marsh edge often collapses abruptly rather than adjusting gradually.
This boundary holds only as long as water can move through it.
Black, organic-rich peat exposed after storms marks the remains of an ancient salt marsh once buried beneath barrier sands. Its reappearance along North Topsail Beach records long-term shoreline change and marsh migration. Photo credit: Bill Tresnan, 2024.
A marsh built by interactions
Across all elevations in Onslow County marshes, the pattern is consistent:
Plants define the zones—but animals and microbes determine whether those zones endure.
Conceptual diagram of revised juvenile blue crab ontogenetic habitat shifts. Arrows depict transitions between habitats with increases in size. Arrow widths denote abundance contributions of individuals between habitats. | Image credit: Hyman et al., 2023
From grazers that keep seagrass blades clean, to crabs that hold the marsh edge together, to microbes that manage invisible chemical stress, the marsh is sustained by small organisms with outsized influence. Together, these interactions determine not just what lives in the marsh, but whether the marsh itself endures.
Purple marsh crabs (Sesarma reticulatum) moving together along the marsh edge on South Topsail Island, North Carolina. Their collective movement and feeding activity illustrate how small organisms play outsized roles in maintaining marsh structure. Photo credit: A. Mitchell, 2025.
References
Abbott, K. M., Quirk, T., & Fultz, L. M. (2022). Soil microbial community development across a 32-year coastal wetland restoration time series and the relative importance of environmental factors. Science of The Total Environment, 821, 153359. https://doi.org/10.1016/j.scitotenv.2022.153359
Bertness, M. D. (1984). Ribbed mussels and Spartina Alterniflora production in a New England salt marsh. Ecology, 65(6), 1794-1807. https://doi.org/10.2307/1937776
Bertness, M. D. (1985). Fiddler crab regulation of Spartina alterniflora production on a New England salt marsh. Ecology, 66(3), 1042-1055. https://doi.org/10.2307/1940564
Duffy, J. E., Reynolds, P. L., Boström, C., Coyer, J. A., Cusson, M., Donadi, S., Douglass, J. G., Eklöf, J. S., Engelen, A. H., Eriksson, B. K., Fredriksen, S., Gamfeldt, L., Gustafsson, C., Hoarau, G., Hori, M., Hovel, K., Iken, K., Lefcheck, J. S., Moksnes, P., … Stachowicz, J. J. (2015). Biodiversity mediates top–down control in eelgrass ecosystems: A global comparative‐experimental approach. Ecology Letters, 18(7), 696-705. https://doi.org/10.1111/ele.12448
Howarth, R. W., & Giblin, A. (1983). Sulfate reduction in the salt marshes at Sapelo island, Georgia. Limnology and Oceanography, 28(1), 70-82. https://doi.org/10.4319/lo.1983.28.1.0070
Hyman, A. C., Chiu, G. S., Seebo, M. S., Smith, A., Saluta, G. G., Knick, K. E., & Lipcius, R. N. (2023). Model-based evaluation of critical nursery habitats for juvenile blue crabs through ontogeny: Abundance and survival in seagrass, salt marsh, and unstructured bottom. https://doi.org/10.1101/2023.07.20.549877
Jordan, T. E., & Valiela, I. (1982). A nitrogen budget of the ribbed mussel, Geukensia demissa, and its significance in nitrogen flow in a New England salt marsh. Limnology and Oceanography, 27(1), 75-90. https://doi.org/10.4319/lo.1982.27.1.0075
Mallin, M. A., Burkholder, J. M., Cahoon, L. B., & Posey, M. H. (2000). North and South Carolina coasts. Marine Pollution Bulletin, 41(1-6), 56-75. https://doi.org/10.1016/s0025-326x(00)00102-8
Mann, K. H. (1988). Production and use of detritus in various freshwater, estuarine, and coastal marine ecosystems. Limnology and Oceanography, 33(4part2), 910-930. https://doi.org/10.4319/lo.1988.33.4part2.0910
Orth, R. J., Heck, K. L., & Van Montfrans, J. (1984). Faunal communities in seagrass beds: A review of the influence of plant structure and prey characteristics on predator: Prey relationships. Estuaries, 7(4), 339. https://doi.org/10.2307/1351618
Orth, R. J., & Van Montfrans, J. (1984). Epiphyte-seagrass relationships with an emphasis on the role of micrograzing: A review. Aquatic Botany, 18(1-2), 43-69. https://doi.org/10.1016/0304-3770(84)90080-9
Silliman, B. R., Van de Koppel, J., Bertness, M. D., Stanton, L. E., & Mendelssohn, I. A. (2005). Drought, snails, and large-scale die-off of southern U.S. salt marshes. Science, 310(5755), 1803-1806. https://doi.org/10.1126/science.1118229
Silliman, B. R., & Zieman, J. C. (2001). Top-down control of Spartina alterniflora production by periwinkle grazing in a Virginia salt marsh. Ecology, 82(10), 2830. https://doi.org/10.2307/2679964
Thomas, C., & Blum, L. (2010). Importance of the fiddler crab Uca pugnax to salt marsh soil organic matter accumulation. Marine Ecology Progress Series, 414, 167-177. https://doi.org/10.3354/meps08708
Valentine, J. F., & Duffy, J. E. (n.d.). The central role of grazing in seagrass ecology. Seagrasses: Biology, Ecology and Conservation, 463-501. https://doi.org/10.1007/1-4020-2983-7_20
Zhang, G., Bai, J., Jia, J., Wang, W., Wang, D., Zhao, Q., Wang, C., & Chen, G. (2023). Soil microbial communities regulate the threshold effect of salinity stress on SOM decomposition in coastal salt marshes. Fundamental Research, 3(6), 868-879. https://doi.org/10.1016/j.fmre.2023.02.024